Доклад: Естественный отбор
Групповой отбор часто называют также групповым отбором, представляет собой дифференциальное размножение разных локальных популяций. В.Райт сравнивает популяционные системы двух типов — большую непрерывную популяцию и ряд мелких полуизолированных колоний — в отношении теоретической эффективности отбора. Предполагается, что общая величина обеих популяционных систем одинакова и организмы свободно скрещиваются между собой.
В большой непрерывной популяции отбор относительно неэффективен в смысле повышения частоты благоприятных, но редких рецессивных мутаций. Кроме того, любой тенденции к повышению частоты какого-либо благоприятного аллеля в одной части данной обширной популяции противодействует скрещивание с соседними субпопуляциями, в которых этот аллель редок. Подобным же образом благоприятные новые генные сочетания, которым удалось образоваться в какой-нибудь локальной доле данной популяции, разбиваются на части и элиминируются в результате скрещивания с особями соседних долей.
Все эти трудности в значительной степени устраняются в популяционной системе, по своей структуре напоминающей ряд отдельных островков. Здесь отбор или отбор совместно с дрейфом генов может быстро и эффективно повысить частоту какого-либо редкого благоприятного аллеля в одной или нескольких мелких колониях. Новые благоприятные сочетания генов также могут легко закрепиться в одной или нескольких мелких колониях. Изоляция защищает генофонды этих колонии от «затопления» в результате миграции из других, не обладающих столь благоприятными генами колоний, и от скрещивания с ними. До этого момента в модель был включен только индивидуальный отбор или — для некоторых колоний — индивидуальный отбор в сочетании с дрейфом генов.
Допустим теперь, что среда, в которой находится данная популяционная система, изменилась, в результате чего адаптивность прежних генотипов понизилась. В новой среде новые благоприятные гены или сочетания генов, закрепившиеся в некоторых колониях, обладают высокой потенциальной адаптивной ценностью для популяционной системы в целом. Теперь имеются все условия для того, чтобы вступил в действие групповой отбор. Менее приспособленные колонии постепенно сокращаются и вымирают, а колонии, которые более приспособлены, расширяются и замещают их по всей области, занимаемой данной популяционной системой. Такая подразделившаяся популяционная система приобретает новый набор приспособительных признаков в результате индивидуального отбора в пределах некоторых колоний, за которым следует дифференциальное размножение разных колоний. Сочетание группового и индивидуального отбора может привести к результатам, которые не могут быть достигнуты за счет одного лишь индивидуального отбора.
Установлено, что групповой отбор — процесс второго порядка, дополняющий главный процесс индивидуального отбора. Будучи процессом второго порядка, групповой отбор должен протекать медленно, вероятно, гораздо медленнее, чем индивидуальный отбор. Обновление популяций требует больше времени, чем обновление особей.
Концепция группового отбора встретила широкое признание в некоторых кругах, но была отвергнута другими учеными.. Они утверждают, что различные возможные модели индивидуального отбора способны вызывать все эффекты, приписываемые групповому отбору. Уэйд провел ряд селекционных экспериментов с мучным хрущаком (Tribolium castaneum), с тем чтобы выяснить эффективность группового отбора, и обнаружил, что жуки реагировали на отбор этого типа. Кроме того, когда на какой-либо признак одновременно действуют индивидуальный и групповой отбор и притом в одном и том же направлении, скорость изменения этого признака выше, чем в случае одного только индивидуального отбора (Даже умеренная иммиграция (6 и 12%) не препятствует дифференциации популяций, вызываемой групповым отбором.

Одна из особенностей органического мира, которую трудно объяснить на основе индивидуального отбора, но можно рассматривать как результат группового отбора, — это половое размножение. Хотя и были созданы модели, в которых половому размножению благоприятствует индивидуальный отбор, однако они представляются нереалистичными. Половое размножение — это тот процесс, который создаёт рекомбинационную изменчивость в скрещивающихся популяциях. От полового размножения выигрывают не родительские генотипы, распадающиеся в процессе рекомбинации, а популяция будущих поколений, у которых возрастает запас изменчивости. Это подразумевает участие в качестве одного из факторов селективного процесса на популяционном уровне.
г) Направленный отбор (движущий)
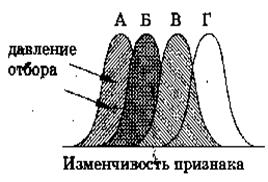
Рис. 1. Движущая форма естественного отбора
Направленный отбор (движущий) был описан еще Ч.Дарвином, а современное учение о движущем отборе разработано Дж. Симпсоном.
Суть этой формы отбора заключается в том, что он вызывает прогрессивное или направленное в одну сторону изменение генетического состава популяций, что проявляется в сдвиге средних значений отбираемых признаков в сторону их усиления или ослабления. Он происходит в тех случаях, когда популяция находится в процессе приспособления к новой среде или же когда происходит постепенное изменение среды, а вслед за ней и постепенное изменение популяции.
При длительном изменении внешней среды преимущество в жизнедеятельности и размножении может получить часть особей вида с некоторыми отклонениями от средней нормы. Это приведет к изменению генетической структуры, возникновению эволюционно новых приспособлений и перестройке организации вида. Вариационная кривая смещается в направлении приспособления к новым условиям существования.
Такой отбор приводит к появлению новой нормы вместо старой, переставшей соответствовать новым условиям. Изменение признака может происходить как в сторону его усиления, так и в сторону ослабления. Как приобретение нового признака, так и утрата какого-либо признака – результаты действия движущей формы отбора. Редукция органов, потерявших свое функциональное значение, происходит именно таким путем. Примерами являются некоторые бескрылые птицы и насекомые, растения-паразиты без корней и листьев, ленточные черви без пищеварительной системы, потемнение окраски бабочки березовой пяденицы в развитых индустриальных районах Англии. До 1848 года описанные в Англии формы бабочек березовой пяденицы имели бледно-кремовую окраску с черными точками и отдельными темными пятнышками. В 1848 году в Манчестере была обнаружена черная форма этой бабочки, а к 1895 году популяция березовой пяденицы в Манчестере состояла на 98% из темноокрашенных бабочек. Эта «меланическая» форма появилась в результате возникновения случайных мутаций, причем фенотип мутантных особей обладал в промышленных районах большим преимуществом по сравнению со светлоокрашенными формами.
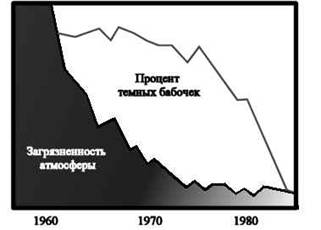
Рис 2.Зависимость частоты темных форм березовой пяденицы от степени загрязненности атмосферы
Светлоокрашенные формы были незаметны на стволах берез, покрытых лишайниками. С интенсивным развитием промышленности диоксид серы, образующийся при сжигании угля, вызывал гибель лишайников в промышленных районах, и в результате обнаружилась темная кора деревьев. На темном фоне светлоокрашенные пяденицы склевывались малиновками и дроздами, а выживали и успешно размножались меланические формы, которые на темном фоне менее заметны. За последние 100 лет у более чем 80 видов бабочек появились темные формы. Это явление известно теперь под названием индустриального (промышленного) меланизма. Движущий отбор приводит к появлению нового вида.


Рис. 3. Индустриальный меланизм. Темные формы бабочек незаметны на темных стволах , а светлые – на светлых.
Насекомые, ящерицы и ряд других обитателей травы имеют зеленую или бурую окраску, обитатели пустыни – цвет песка. Шерсть животных, живущих в лесах, например леопарда, расцвечена небольшими пятнами, напоминающими солнечные блики, а у тигра имитирует окраску и тень от стеблей камыша или тростника. Такая окраска получила название покровительственной.
У хищников она закрепилась благодаря тому, что ее обладатели незаметно могли подкрадываться к добыче, а у организмов, являющихся добычей, - вследствие того, что жертва оставалась менее заметной для хищников. Как же она появилась? Многочисленные мутации давали и дают большое разнообразие форм, отличающихся по окраске. В ряде случаев расцветка животного оказывалась близкой к фону окружающей среды, т.е. скрывала животное, играла роль покровительственной. Те животные, у которых покровительственная окраска была выражена слабо, оставались без пищи либо сами становились жертвой, а их сородичи, обладающие лучшей покровительственной окраской, выходили победителями в межвидовой борьбе за существование.
Направленный отбор лежит в основе искусственного отбора, при котором избирательное скрещивание особей, обладающих желательными фенотипическими признаками, повышает частоту этих признаков в популяции. В ряде экспериментов Фальконер выбирал из популяции шестинедельных мышей самых тяжелых особей и давал им спариваться между собой. То же самое он проделывал с самыми легкими мышами. Такое избирательное скрещивание по признаку массы тела привело к созданию двух популяций, в одной из которых масса возрастала, а в другой – уменьшалась.
После прекращения селекции ни та, ни другая группа не вернулась к первоначальной массе (примерно 22 грамма). Это показывает, что искусственный отбор по фенотипическим признакам привел к некоторому генотипическому отбору и частичной утрате обеими популяциями каких-то аллелей.
д) Стабилизирующий отбор
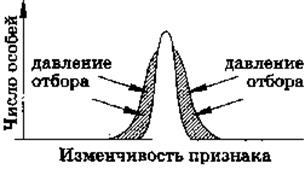
Рис. 4. Стабилизирующая форма естественного отбора
Стабилизирующий отбор в относительно постоянных условиях среды естественный отбор направлен против особей, признаки которых отклоняются от средней нормы в ту или другую сторону.
Стабилизирующий отбор сохраняет то состояние популяции, которое обеспечивает ее максимальную приспособленность в постоянных условиях существования. В каждом поколении удаляются особи, отклоняющиеся от среднего оптимального значения по приспособительным признакам.
Описано множество примеров действия стабилизующего отбора в природе. Например, на первый взгляд кажется, что наибольший вклад в генофонд следующего поколения должны вносить особи с максимальной плодовитостью.
