Доклад: Волновые процессы в зрительной коре мозга
Карты альфа-потенциалов (с шагом 4 мс) мы строили на основе ЭЭГ испытуемых, которые находились в условиях наибольшего проявления альфа-ритма, т.е. спокойно сидели закрыв глаза. Движущиеся волны обнаруживали, сравнивая ЭЭГ, полученные до и во время фотостимуляции, и ранжировали их по 10 траекториям. Каковы же были результаты?
Во-первых, на серии динамических карт, отражающих волну, которая двигалась от затылка ко лбу, мы увидели, что отрицательный полюс потенциала, находящийся в затылочной области коры, распространяется в том же направлении [8]. Во-вторых, удалось выявить, что конкретные иллюзии возникали на фоне определенной траектории волны. Например, круг и спираль появлялись в том случае, если волна двигалась от затылка ко лбу, а решетка — при траектории, направленной от левой затылочной области коры к правой лобной. Следовательно, связь между видом иллюзии и направлением движения альфа-волны существует.
Прямое доказательство
Теперь мы уже знали, что от фазы альфа-волны зависит распознавание формы геометрических фигур и направления движения зрительного стимула, что эту волну можно “остановить”, вызвав ее иллюзорный зрительный образ; увидели, что она распространяется по нескольким траекториям, с которыми связан определенный вид иллюзии. Но все это — косвенные доказательства. Как же получить прямые подтверждения теоретическим построениям Питса и Мак-Каллока?
Исходя из зависимости вида иллюзии от траектории смещения максимума потенциала, мы предположили, что эта траектория может отражать и движение источника альфа-ритма. Обнаружив это, мы тем самым непосредственно доказали бы гипотезу сканирования.
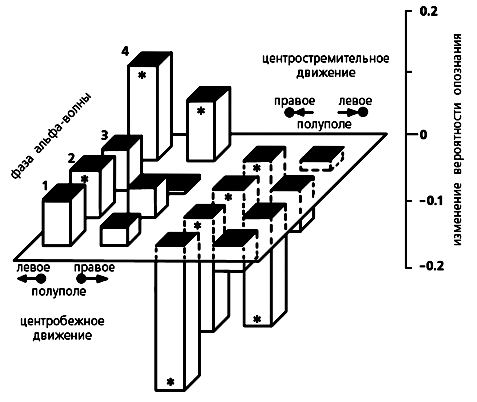
Опознание центробежного и центростремительного движений. При совпадении времени запуска движущегося светового пятна с какой-либо фазой альфа-волны центробежное направление опознается лучше по сравнению с контролем (нулевая плоскость), а центростремительное — хуже. Предпочтение выражено сильнее, когда пятно начинает движение из левого полуполя зрения. Звездочками на схеме отмечены достоверные (р<0.05) отклонения в опознании.
Регистрация электрической активности мозга от набора электродов, расположенных в разных участках головы человека, позволяет судить о распределении потенциалов на двумерной поверхности. Известно, однако, что в энцефалограмму существенный, а иногда и определяющий вклад вносят генераторы потенциалов из глубинных структур мозга. Обнаружить и установить местоположение таких генераторов позволяют методы трехмерного анализа ЭЭГ, основанные на решении обратной задачи. Для этого разработаны специальные компьютерные программы.
Считается, что за распределение альфа-потенциалов по поверхности головы отвечает эквивалентный токовый диполь альфа-ритма [9]. Наша задача состояла в том, чтобы выявить последовательный сдвиг диполя по зрительной коре во время развития альфа-волны, которая регистрируется от поверхности головы.
Для этого анализировали ЭЭГ испытуемых (в опытах с ритмической фотостимуляцией, вызывающей зрительные иллюзии), решая обратную задачу для трехслойной сферической модели головы. В результате в затылочных долях мозга удалось выявить и последовательный сдвиг диполя, и поворот его вектора [10]. При движении альфа-волны от затылочной области к лобной источник альфа-ритма смещался в медиальном направлении и вниз, а его дипольный момент поворачивался против часовой стрелки в горизонтальной плоскости и по часовой стрелке — в сагиттальной. Эти результаты указывают на быстрое смещение источника альфа-ритма в области мозга, которая совпадает с первичной зрительной корой, и подтверждают гипотезу о ее сканировании распространяющейся волной.
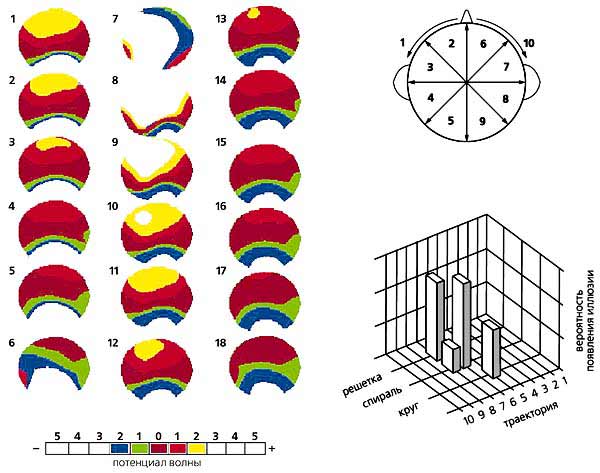
Движущаяся волна альфа-ритма и ее траектории. На картах поля альфа-потенциалов мозга (три левых колонки ) одного из испытуемых видно, что она распространяется от затылочной области мозга к лобной. Возможны несколько траекторий (на правой части рисунка их показано 10). На фоне определенной траектории и возникает конкретная зрительная иллюзия (справа внизу ).
Таким образом, последовательный сдвиг диполя во время альфа-ритма прямо отражает движение волны возбуждения по зрительной коре. Поворот же вектора диполя при этом становится неизбежным, ведь она движется не по ровной поверхности, а по сложно упакованной шпорной извилине в первичной проекционной области зрительной коры.
Поскольку локализация источника альфа-ритма проводилась на “обобщенной” трехмерной модели головы, мы сочли необходимым подтвердить его местоположение для каждого испытуемого с помощью ЯМР-томографии. Полученные трехмерные карты структур мозга (с пространственным разрешением около 1 мм) каждого конкретного человека, участвовавшего в опыте, совмещали по определенной программе с картами, которые отражали локализацию диполей альфа-ритма. Благодаря такому наложению двух видов трехмерных карт впервые удалось прямо выявить, что дипольный источник альфа-волны во время ее движения последовательно смещается по шпорной извилине. Это непосредственно, а не косвенно, свидетельствует в пользу гипотезы сканирования.
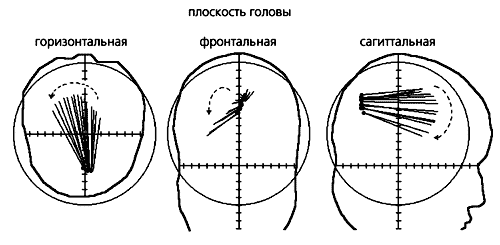
Положение токового диполя альфа-ритма и его векторов в динамике (шаг 2 мс). Токовый диполь (черные точки) — источник альфа-ритма — локализован в затылочной области, в какой бы плоскости профиля головы он ни был выявлен, и его позиция меняется по мере движения волны. Происходит и поворот вектора диполя (линии, исходящие из точек).
Механизмы генерации и распространения альфа-волны
Теперь, имея доказательства гипотезы Питса и Мак-Каллока, попробуем разобраться в механизмах, которые обусловливают возникновение и движение альфа-волны.
В принципе полученные в наших экспериментах эффекты можно было бы попытаться объяснить и без привлечения идеи о движении альфа-волны по зрительной коре. Однако если предположить, что одновременно вся эта кора просто “дышит” с частотой альфа-ритма, то нельзя было бы связать зависимость опознания геометрических фигур разного размера с фазой альфа-волны. Необъяснимым осталось бы и появление оформленных зрительных иллюзий при фотовспышках с альфа-частотой.
Часто критики гипотезы сканирования справедливо напоминают, что альфа-ритм наиболее выражен в темноте и при закрытых глазах, а воздействие света подавляет его, и он сменяется асинхронными колебаниями на более высоких частотах. Но противоречие этого известного факта с гипотезой Питса и Мак-Каллока только кажущееся, и авторы, кстати, сами упоминали о нем. Можно думать, что активация альфа-ритма при закрытых глазах — это эффект “холостого хода” сканирующего механизма. Тогда снижение на свету мощности альфа-ритма свидетельствует не об его истинном уменьшении, а об интерференционной маскировке. В самом деле, пришедшие из сетчатки глаза сигналы о многообразном внешнем мире по-разному активируют корковые нейроны, которыми эти сигналы (точнее, зрительная информация) перерабатываются. В результате создается множество волн возбуждения не синхронных с альфа-ритмом, потому и возникает впечатление, что он затухает при активном зрительном восприятии.
Питс и Мак-Каллок считали, что благодаря сканированию зрительной коры альфа-волной человек распознает объекты внешнего мира независимо от изменения их размера и поворота в пространстве. (Все мы узнаем стул, большой он или маленький, стоит на ногах или лежит на боку.) Кроме того, авторы гипотезы утверждали, что последовательное сканирование выгодно для представления на выходе зрительной коры информации не только в пространственном коде, но и во временном. К тому же сканирование, считали они, может обеспечить компактность “кабеля”, который связывает первичную зрительную кору с другими корковыми полями мозга. И этот “кабель” не требует того громадного числа нервных волокон, которое было бы необходимо для восприятия зрительной информации без сканирующего процесса. Благодаря ему, утверждали Питс и Мак-Каллок, такой “кабель” может быть упакован в отведенное ему ограниченное место в белом веществе затылочных долей мозга. Трудно добавить что-либо новое и существенное к этим весьма развитым умозрительным построениям, высказанным 50 лет назад. И хотя их все еще трудно обсуждать в деталях, аналогии можно найти. Отечественный физиолог М.Н.Ливанов (1907—1986) говорил о пользе сонастройки ритмов активности разных структур мозга для обеспечения функциональной связи между ними. За счет такой связи и повышается эффективность двигательной реакции на сенсорный стимул у животных и человека.
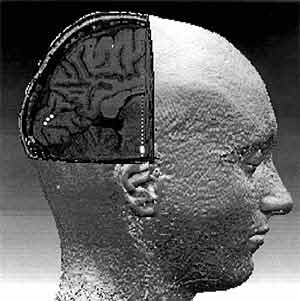
Точная локализация токовых диполей в мозге человека. На трехмерной карте мозга, полученной по данным ЯМР-томографии, расчетным методом виртуальной хирургии удалена часть затылочного полюса правого полушария. На внутренней, “обнажившейся” поверхности левого полушария показаны три последовательных положения дипольного источника альфа-ритма (белые точки). Находятся они на шпорной борозде — первичной проекционной области зрительной коры (другие дипольные источники располагаются глубже и на срезе не видны). Из-за сложности рельефа этой борозды поворот вектора диполя становится неизбежным.
Известно, что волны возбуждения могут генерироваться как внутри коры мозга, так и в подкорке, т.е. в структурах, внешних по отношению к ней. В самой коре волны, движущиеся со скоростью нескольких метров в секунду (такова скорость и сканирующей волны), до сих пор прямо не визуализированы. Правда, благодаря применению все более совершенных технических устройств в нейрофизиологических исследованиях уже удалось увидеть более медленные, чем сканирующая альфа-волна, волны (со скоростями до нескольких десятков мм/c) в зрительной коре [11] и обонятельной луковице [12]. Мы надеемся, что удастся увидеть, а не иллюзорно воспринять, и движущуюся альфа-волну. Мы уверены, что ее визуализация позволит лучше понять последовательность операций по переработке зрительной информации в коре большого мозга.
Литература
1. Pitts W., McCulloch W.S. // Bull. Math. Biophys. 1947. V.9. P.127—147.
2. Шевелев И.А. и др. // Физиология человека. 1985. Т.11. №5. С.707—711.
3. Shevelev I.A. et al. // Int. J. Psychophysiol. 1991. V.11. P.195—201.
4. Шевелев И.А. и др. // Сенсор. системы. 1991. Т.5. №3. С.54—59.