Контрольная работа: Синапсы. Центральная нервная система
Нейрон имеет дендриты, аксон, сому. Для физиолога очень важно понятие об аксонном холмике и начальном сегменте. Именно в этом месте происходит возбуждение нейрона, так как эта часть обладает наибольшей возбудимостью.
Нейрон имеет множество синапсов, через которые он получает возбуждение и тормозные воздействия от других нейронов. Благодаря этому нейрон может получать в больших количествах информацию.
Нейрон может находиться в различных состояниях: а) в состоянии покоя – практически отсутствуют колебания мембранного потенциала, ПД не генерируется; б) в состоянии активности – генерировать потенциалы действия (для нейронов характерна генерация серии или пачки импульсов). Состояние активности может быть индуцировано за счет поступления к нейрону импульсов от других нейронов или быть спонтанным (автоматия). В этом случае нейрон играет роль пейсмекера (водителя ритма). Такие нейроны имеются в ряде центров, например, в центре дыхания; в) в состоянии торможения – оно проявляется в том, что нейрон прекращает свою импульсную активность (нейрон – пейсмекер, или нейрон, получающий возбуждающие воздействия). В основе торможения лежит явление гиперполяризации нейрона (это характерно для постсинаптического торможения).
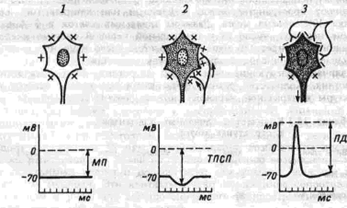 |
Возможные состояния нейрона1 – поляризация, 2 – гиперполяризация, 3 – деполяризация; МП – мембранный потенциал, ТПСП – тормозящий постсинаптический потенциал, ПД – потенциал действия |
В нервной клетке при развитии потенциала действия возникает период рефрактерности или невозбудимости. Это явление заключается в том, что на фоне потенциала действия и еще некоторое время после его завершения, нельзя вызвать второй разряд клетки, невзирая на силу применяемого раздражения. Время, в течение которого нервная клетка невозбудима, называется абсолютной рефрактерностью. Затем клетка приобретает возможность реагировать на прилагаемые раздражения, но впервые возникающий после периода абсолютной рефрактерности потенциал имеет меньшую амплитуду, так как возникает при меньшем уровне заряда мембраны. Периоды появления неполных ответов называются периодами относительной рефрактерности.
Функционально нейроны делят на три типа: афферентные, промежуточные и эфферентные. Первые – выполняют функцию получения и передачи информации в вышележащие структуры ЦНС, вторые – обеспечивают взаимодействие между нейронами одной структуры, третьи – за счет длинного аксона передают информацию в нижележащие структуры ЦНС, в нервные узлы, лежащие за ее пределами, и в органы организма.
По форме нейроны делят на моно-, би- и мультиполярные. По химической характеристике выделяемых в окончаниях аксонов веществ, отличают нейроны: холинэргические, пептидэргические, норадреналинэргические, дофаминэргические, серотонинэргические и др.
Важной характеристикой нейронов является их чувствительность к разным раздражителям. По этому признаку нейроны делят на моно-, би- и полисенсорные.
Моносенсорные нейроны располагаются чаще в первичных проекционных зонах коры и реагируют только на сигналы своей модальности. Например, значительная часть нейронов первичной зрительной коры реагирует только на световое раздражение сетчатки глаза.
Бисенсорные нейроны располагаются преимущественно во вторичных зонах коры анализатора и могут реагировать как на сигналы своей, так и на сигналы другой модальности. Например, нейроны вторичной зрительной коры реагируют на зрительные и слуховые раздражения. Полисенсорные нейроны – это чаще всего нейроны ассоциативных зон мозга. Они способны реагировать на раздражение слуховой, зрительной, кожной и др. анализаторных систем.
Функции нейроглии. Глия – структура нервной системы, образованная специализированными клетками различной формы, которые заполняют пространства между нейронами или капиллярами, составляя 10% объема мозга. Размеры глиальных клеток в 3–4 раза меньше нервных, число их в центральной нервной системе млекопитающих достигает 140 млрд. С возрастом число нейронов в мозгу уменьшается, а число глиальных клеток увеличивается.
Различают следующие виды глии: астроглия, олигодендроглия, микроглия.
Астроглия – представлена многоотростчатыми клетками. Их размеры колеблются от 7 до 25 мкм. Большая часть отростков заканчивается на стенках сосудов. Астроглия служит опорой нейронов, обеспечивает репаративные процессы нервных стволов, изолирует нервное волокно, участвует в метаболизме нейронов.
Олигодендроглия – это клетки, имеющие один отросток. Количество олигодендроглии возрастает в коре от верхних слоев к нижним. В подкорковых структурах, в стволе мозга олигодендроглии больше, чем в коре. Она участвует в миелинизации аксонов, в метаболизме нейронов.
Микроглия – самые мелкие клетки глии, относятся к блуждающим клеткам. Они образуются из структур оболочек мозга, проникают в белое, а затем и в серое вещество мозга. Микроглиальные клетки способны к фагоцитозу.
Одной из особенностей глиальных клеток является их способность к изменению своего размера. Изменение размера глиальных клеток носит ритмический характер: фазы сокращения – 90 с, расслабления – 240 с, т.е. это очень медленный процесс. Средняя частота ритмических изменений варьирует от 2 до 20 в час. При этом отростки клетки набухают, но не укорачиваются в длине.
Физиологические процессы в нервной системе во многом зависят от миелинизации волокон нервных клеток. В центральной нервной системе миелинизация обеспечивается олигодендроглией, а в периферической – шванновскими клетками.
Торможение в ЦНС его виды и предполагаемые механизмы
Впервые идею о том, что в ЦНС помимо процессов возбуждения существует процесс торможения, высказал И.М. Сеченов.
Если рассмотреть «архитектуру» использования тормозных нейронов при организации нейронных сетей, цепей и рефлекторных дуг, то можно выделить ряд вариантов этой организации (это отражается в названии данного вида торможения).
1. Реципрокное торможение. Как пример: сигнал от мышечного веретена поступает с афферентного нейрона в спинной мозг, где переключается на альфа-мотонейрон сгибателя и одновременно на тормозной нейрон, который тормозит активность альфа-мотонейрона разгибателя. Явление открыто Ч. Шеррингтоном.
2. Возвратное торможение. Альфа-мотонейрон, к примеру, посылает аксон к соответствующим мышечным волокнам. По пути от аксона отходит коллатераль, которая возвращается в ЦНС – она заканчивается на тормозном нейроне (клетка Реншоу) и активирует ее. Тормозной нейрон вызывает торможение альфа-мотонейрона, который запустил всю эту цепочку. Таким образом, альфа-мотонейрон, активируясь, через систему тормозного нейрона сам себя тормозит.
3. Существует ряд вариантов возвратного торможения, в частности, латеральное торможение. Суть его сводится к тому, что активируется, например, фоторецептор, он активизирует биполярную клетку, и одновременно активируется рядом расположенный тормозной нейрон, который блокирует проведение возбуждения от соседнего фоторецептора к ганглиозной клетке. Этим самым происходит «вытормаживание» информации в соседних участках. Таким способом создаются условия для четкого видения предмета (две точки на сетчатке рассматриваются как две раздельные точки в том случае, если между ними есть невозбужденные участки).

Особенности распространения возбуждения в ЦНС
Рассматривая основные принципы и особенности распространения возбуждения в ЦНС, а по сути, и свойства нервных центров, можно выделить следующие моменты.
1. Одностороннее проведение возбуждения. В ЦНС – в ее центрах, внутри рефлекторной дуги и нейронных цепей возбуждение, как правило, идет в одном направлении, например, от афферентного нейрона к эфферентному, а не наоборот. Обусловлено это особенностями расположения и характером функционирования химического синапса.