Курсовая работа: Нервная ткань
В отличие от непостоянного количества дендритов у любой нервной клетки может быть только один аксон, который проводит электрические сигналы лишь в одном направлении: от тела клетки. Эти электрические сигналы называются потенциалами действия и обычно возникают в аксонном холмике - месте отхождения аксона от сомы и распространяются по аксону со скоростью от 1 до 100 м/с.
Диаметр аксона почти одинаков на всем протяжении, у разных клеток его величина варьирует от 0,2 до 20 мкм. Это обстоятельство сказывается на скорости проведения сигнала: чем толще аксон, тем быстрее проводится по нему потенциал действия. Длина аксонов у разных клеток может сильно отличаться: от 0,1 мм до 1м. Многие аксоны заключены в особый футляр, сформированный отростками некоторых клеток глии. Образует этот футляр миелин - жироподобное вещество со свойствами электрического изолятора: в области миелиновых покрытий электрические сигналы не возникают.
Миелиновая оболочка аксона регулярно прерывается участками, свободными от миелина - они называются перехватами Ранвье. По этим перехватам и распространяются потенциалы действия, как бы перескакивая через миелинизированные участки от одного перехвата к другому (такой тип передачи называется сальтаторным, от лат. saltare - прыгать), поэтому скорость проведения оказывается довольно высокой. Некоторые аксоны не имеют миелинового покрытия: в отличие от миелинизированных волокон их называют безмиелиновыми (по другой терминологии миелинизированные и безмиелиновые волокна различают как мякотные и безмякотные). По безмиелиновым волокнам потенциалы действия распространяется медленнее: здесь они не «прыгают», а «ползут» по всей длине аксона.
Неподалеку от своих окончаний большинство аксонов разделяется на тонкие коллатеральные ветви или аксонные терминали, причем некоторые из них могут повернуть назад - это обратные коллатерали. Терминали аксона вступают в контакт с другими клетками, чаще всего с их дендритами, реже - с телом и еще реже - с аксоном. Аксоны эфферентных нейронов контактируют с клетками рабочих органов, которыми являются мышцы или железы внешней секреции. Контактная зона между двумя клетками получила название: синапс. В соответствии с этим термином клетка, передающая сигнал, называется пресинаптической, а получающая сигнал- постсинаптической. В подавляющем большинстве случаев эти клетки анатомически не соединяются и между ними находится синаптическая щель, которая заполнена жидкостью, напоминающей по своему составу плазму крови.
Из-за анатомической разобщенности пресинаптическая клетка может повлиять на постсинаптическую только с помощью химического посредника нейромедиатора или нейротрансмиттера. Медиатор должен выделиться из окончания аксона пресинаптической клетки тогда, когда к этому окончанию подойдет потенциал действия.
Классификация нейронов
По количеству цитоплазматических отростков принято различать униполярные, биполярные и мультиполярные нейроны. Униполярные нейроны имеют единственный, обычно сильно разветвленный первичный отросток. Одна из его ветвей функционирует как аксон, а остальные - как дендриты. Такие клетки часто встречаются в нервной системе беспозвоночных, а у позвоночных они обнаруживаются лишь в некоторых ганглиях вегетативной нервной системы.
У биполярных клеток есть два отростка: дендрит проводит сигналы от периферии к телу клетки, а аксон передает информацию от тела клетки к другим нейронам. Так выглядят, например, некоторые сенсорные нейроны, встречающиеся в сетчатке глаза, в обонятельном эпителии. К этой же разновидности нейронов следует отнести и чувствительные клетки спинальных ганглиев, воспринимающих, например, прикосновение к коже или боль, хотя формально от их тела отходит лишь один отросток, который разделяется на центральную и периферическую ветви. Такие клетки называют псевдоуниполярными, они формировались первоначально как биполярные нейроны, но в процессе развития два их отростка соединились в один, у которого одна ветвь функционирует как аксон, а другая - как дендрит.
Типы нейронов
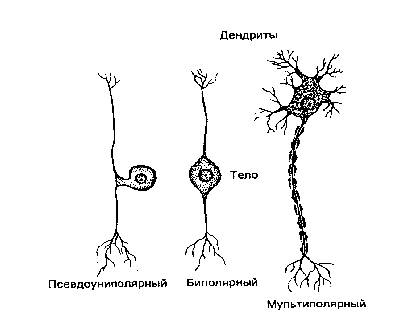
У мультиполярных клеток один аксон, а дендритов может быть очень много, они отходят от тела клетки, а затем многократно делятся, образуя на своих ветвях многочисленные синапсы с другими нейронами. Так, например, на дендритах только одного мотонейрона спинного мозга образуется около 8000 синапсов, а на дендритах находящихся в коре мозжечка клеток Пуркинье может быть до 150000 синапсов.
Нейроны Пуркинье и самыми крупными клетками человеческого мозга: диаметр их тела около 80 мкм. А рядом с ними обнаруживаются крохотные зернистые клетки, их диаметр всего лишь 6-8 мкм. Мультиполярные нейроны встречаются в нервной системе чаще всего и среди них выявляется множество внешне не похожих друг на друга клеток. Нейроны принято классифицировать не только по форме, но и по выполняемой функции, по их месту в цепи взаимодействующих клеток. Некоторые из них имеют специальные чувствительные окончания - рецепторы, которые возбуждаются при действии на них каких-либо физических или химических факторов, таких как, например, свет, давление, присоединение определенных молекул. После возбуждения рецепторов чувствительные нейроны передают информацию в центральную нервную систему, т. е. проводят сигналы центростремительно или афферентно (лат. afferens - приносящий).
Другая разновидность клеток передает команды от центральной нервной системы к скелетным или гладким мышцам, к сердечной мышце или железам внешней секреции. Это либо двигательные, либо вегетативные нейроны, по которым сигналы распространяются центробежно, а сами такие нейроны называются эфферентными (лат. efferens - выносящий).
Все остальные нейроны относятся к категории вставочных или интернейронов, которые образуют основную массу нервной системы - 99,98% от общего количества клеток. Среди них встречаются локальные и проекционные нейроны. Другое название проекционных нейронов - релейные; у них, как правило, длинные аксоны, с помощью которых эти клетки могут передавать переработанную информацию отдаленным регионам мозга. У локальных интернейронов аксоны короткие, эти клетки перерабатывают информацию в ограниченных локальных цепях и взаимодействуют преимущественно с соседними нейронами.
Электрические сигналы
Современный вариант нейронной теории связывает определенные части нервной клетки с характером возникающих в них электрических сигналов. В типичном нейроне есть четыре определяемые морфологические области: сома, дендриты, аксон и пресинаптическое окончание аксона. При возбуждении нейрона в нем последовательно появляется четыре разновидности электрических сигналов: входной, объединенный, проводящийся и выходной. Каждый из этих сигналов возникает только в определенной морфологической области.
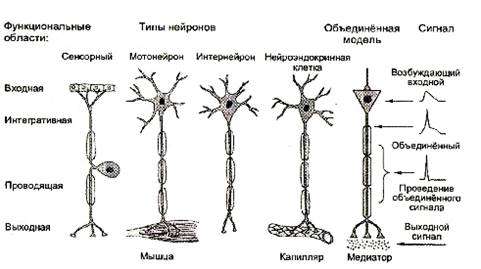
Четыре функциональные области нейронов
Чтобы понять различия между этими сигналами, надо иметь некоторое представление о природе нервных импульсов. С наружной и внутренней стороны плазматической мембраны нейрона содержатся разные электрические заряды: с наружной с тороны положительные, с внутренней - отрицательные. Разность между ними называется мембранным потенциалом покоя. Если считать наружный заряд равным нулю, то разность зарядов между наружной и внутренней поверхностями у большинства нейронов оказывается близкой к 65мВ, хотя она и может у отдельных клеток варьировать от - 40 до -80мВ.
Возникновение этой разности зарядов обусловлено неодинаковым распределением ионов калия, натрия и хлора внутри клетки и снаружи ее, а также большей проницаемостью покоящейся клеточной мембраны лишь для ионов калия.
У возбудимых клеток, к числу которых принадлежат нервные и мышечные, потенциал покоя способен сильно изменяться и эта способность является основой для возникновения электрических сигналов. Уменьшение потенциала покоя, например, с -65 до -60мВ, называется деполяризацией, а увеличение, например, с -65 до -70мВ, - гиперполяризацией.
Если деполяризация достигнет некоторого критического уровня, например -55мВ, то проницаемость мембраны для ионов натрия на короткое время становится максимальной, они устремляются в клетку и в связи с этим трансмембранная разность потенциалов стремительно уменьшается до 0, а затем приобретает положительное значение. Это обстоятельство приводит к закрытию натриевых каналов и стремительному выходу из клетки ионов калия через предназначенные только для них каналы: в результате восстанавливается первоначальная величина мембранного потенциала. Эти быстро происходящие изменения мембранного потенциала называются потенциалом действия. Потенциал действия является проводящимся электрическим сигналом, он быстро распространяется по мембране аксона до самого его окончания, причем нигде не меняет свою амплитуду.
Кроме потенциалов действия в нервной клетке, вследствие изменения ее мембранной проницаемости, могут возникать местные или локальные сигналы: рецепторный потенциал и постсинаптический потенциал. Их амплитуда значительно меньше, чем у потенциала действия, кроме того, она существенно уменьшается при распространении сигнала. По этой причине местные потенциалы и не могут распространяться по мембране далеко от места своего возникновения.
Входные сигналы
Входными сигналами являются либо рецепторный, либо постсинаптический потенциал. Рецепторный потенциал образуется в окончаниях чувствительного нейрона, когда на них действует определенный стимул: растяжение, давление, свет, химическое вещество и т. п. Действие стимула вызывает открытие определенных ионных каналов мембраны, а последующий ток ионов через эти каналы изменяет первоначальное значение потенциала покоя; в большинстве случаев происходит деполяризация. Эта деполяризация и является рецепторным потенциалом, ее амплитуда пропорциональна силе действующего стимула. Рецепторный потенциал может распространяться от места действия стимула вдоль мембраны, но на относительно небольшое расстояние.
Вторая разновидность входного потенциала - постсинаптический потенциал. Он образуется на постсинаптической клетке после того, как возбужденная пресинаптическая отправит для нее специального химического курьера - нейромедиатор. Добравшись путем диффузии до постсинаптической клетки, медиатор присоединяется к рецепторам ее мембраны, что вызывает открытие ионных каналов. Возникший в связи с этим ток ионов через постсинаптическую мембрану изменяет первоначальное значение потенциала покоя - этот сдвиг и является постсинаптическим потенциалом.
В одних синапсах такой сдвиг представляет собой деполяризацию и, если она достигнет критического уровня, то постсинаптический нейрон возбуждается. В других синапсах возникает противоположный по направленности сдвиг: постсинаптическая мембрана гиперполяризуется: величина мембранного потенциала становится больше и уменьшить ее до критического уровня деполяризации становится труднее. Такую клетку трудно возбудить, она заторможена. Таким образом, деполяризующий постсинаптический потенциал является возбуждающим а гиперполяризующий - тормозным. Соответственно этому и сами синапсы подразделяются на возбуждающие (вызывающие деполяризацию) и тормозные (вызывающие гиперполяризацию). Возбуждающие синапсы образуются, по большей части на дендритах постсинаптической клетки, а тормозные - на ее теле.
Вне зависимости от того, что происходит на постсинаптической мембране: деполяризация или гиперполяризация, величина постсинаптических потенциалов всегда пропорциональна количеству подействовавших молекул медиатора, но обычно их амплитуда невелика. Так же, как и рецепторный потенциал, они распространяются вдоль мембраны на очень небольшое расстояние, т. е. относятся к местным потенциалам.
Таким образом, входные сигналы представлены двумя разновидностями местных потенциалов, рецепторным и постсинаптическим, а возникают эти потенциалы в строго определенных областях нейрона: либо в чувствительных окончаниях, либо в синапсах. Чувствительные окончания принадлежат сенсорным нейронам, где рецепторный потенциал возникает под действием каких-либо раздражителей, внешних по отношению к нейрону стимулов. Для интернейронов, а также для эфферентных нейронов входным сигналом может быть только постсинаптический потенциал.