Курсовая работа: Строение и принцип действия переносчиков
Первичные активные переносчики генерируют напряжение и создают трансмембранные ионные градиенты. Вторичные активные переносчики используют такие градиенты в качестве движущей силы для транспорта растворимых веществ. Наиболее полно охарактеризованным примером такого рода является белок - переносчик лактозы (лактозопермеаза) из Escherichia colt. Этот переносчик использует протонный электрохимический градиент, генерируемый дыхательной электронтранспортной цепью, в качестве движущей силы для накопления лактозы в клетке. Это пример симпорта, когда через мембрану одновременно переносятся два разных вещества (например, протоны и лактоза). Антипортеры осуществляют транспорт веществ в противоположных направлениях. Так, например, белок полосы 3 эритроцитов осуществляет сопряженный транспорт Cl - и НСО3 - в противоположных направлениях через эритроцитарную мембрану.
Термины пермеаза, транслоказа и переносчик, являющиеся синонимами, их часто используют по отношению к транспортным белкам, отличным от первичных активных переносчиков. Обычно термин "пермеаза" применяют при описании бактериальных транспортных белков. Термин "переносчик", по-видимому, лучше использовать по отношению к ионофорам или сходным с ними структурам, которые связываются с ионами и переносят их через биослой в составе комплекса.
Некоторые группы переносчиков:
1. Митохондриальные переносчики:
переносчик ADP/ATP; - переносчик Н+ - фосфатаза;
разобщающий белок (переносчик Н+ /ОН -)
2. Переносчики сахаров:
переносчик глюкозы (клетки млекопитающих);
переносчик Н+-арабинозы (Е. coli);
переносчик Н+ - ксилозы (Е. coli);
3. АТРазы E1E2-типа, сопряженные с трансмембранным переносом ионов:
Н+ /К+-АТРаза (слизистая желудка млекопитающих);
Na+ /К+ - АТРаза (плазматическая мембрана);
Са2+-АТРаза (саркоплазматический ретикулум);
Н+ - АТРаза (плазматическая мембрана);
К+-АТРаза (S. faecalis).
1. Переносчики: разнообразие функций
Функции переносчиков весьма разнообразны; проиллюстрируем их на нескольких примерах.
Таблица 2. Сравнение скоростей транспорта для некоторых систем.
| Система | Скорость транспорта, с-1 |
| Н+-лактозопермеаза Е | 30 |
| Переносчик глюкозы(эритроциты) | 300 |
| Анионный переносчик белок полосы 3 | 100 000 |
| Бактериородопсин | 50 |
| Nа+/К+-АТРаза | 450 |
| Цитохром с-оксидаза | 1000 |
В табл.2 приведены значения числа оборотов для нескольких переносчиков. Для лактозопермеазы Е. coli максимальное число оборотов составляет всего лишь 30 с-1. Ее роль состоит в транспорте лактозы - углевода, который затем участвует в клеточном метаболизме.
Для ионных насосов, использующих для работы энергию гидролиза АТР или переноса электронов, характерны максимальные числа оборотов, что довольно типично для ферментов.
Однако не все переносчики работают столь медленно. Анионный переносчик белок полосы 3 из эритроцитарной мембраны играет важную физиологическую роль в усилении быстрого трансмембранного обмена С1 - на НСО3-. Одна из функций эритроцитов заключается в усилении транспорта СО2 от различных тканей к легким. В венозных капиллярах СО2 быстро диффундирует через эритроцитарную мембрану. В клетке под действием карбоангидразы СО2 превращается в Н2СО3, затем быстро устанавливается равновесие Н2СО3 ↔ Н+ + НСО3 - , и анион бикарбоната переносится через мембрану в плазму крови белком полосы 3. В результате по мере того, как эритроцит проходит по капиллярам, концентрация НСО3 - в плазме увеличивается, причем этот процесс занимает меньше 1 с. Когда кровь достигает легких, начинается диффузия СО2 в атмосферу. При этом под действием карбоангидразы в эритроцитах происходит массовое превращение Н2СО3 в СО2 и Н2О. Этот процесс в свою очередь является движущей силой для переноса аниона бикарбоната внутрь эритроцита, где он быстро превращается в СО2 и Н2О.
Транспортная система должна функционировать очень быстро, но в отличие от ионных каналов в аксонах здесь нет нужды в электрогенных реакциях, которые только замедлили бы быстрый массовый транспорт. Но транспорт катиона, например Na+, вместе с НСО3 - был бы нежелателен, поскольку изменение концентрации соли в эритроците привело бы к осмотическому дисбалансу. Эта проблема решается с помощью антипортера, который в обмен на каждый транспортируемый ион НСО3 - переносит в обратном направлении анион Сl-. Такая челночная система работает очень быстро.
2. Переносчики как ферменты: применение теории скоростей
Кинетическую теорию переходного состояния Эйринга, используемую энзимологами, успешно применяют и в случае различных транспортных систем. В основе этого подхода лежит предположение о том, что система может находиться в нескольких дискретных состояниях, каждому из которых соответствует стандартное значение электрохимического потенциала. При этом взаимные переходы между двумя состояниями сопряжены с переходом системы через промежуточные стадии с более высокой свобод ной энергией, и константы скоростей переходов зависят от высоты соответствующих энергетических барьеров. Минимумы на кривых изменения свободной энергии (рис.1) соответствуют местам связывания транспортируемых веществ. Можно предположить, что переносчик имеет одно или несколько мест связывания переносимых веществ. При достаточно высоких концентрациях переносимого вещества все эти места оказываются занятыми и скорость переноса достигает своего максимального значения К mах, равного максимальной скорости работы фермента. Экспериментальные подтверждения этому получены для всех переносчиков.
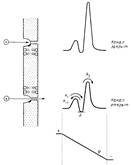
Рис.1
3. Применение теории переходного состояния при изучении работы переносчиков
Рассмотрим простой переносчик с одним местом связывания, транспортирующий молекулы через мембрану. Рис.2 иллюстрирует основные свойства как первичного активного переносчика, так и пермеазы.
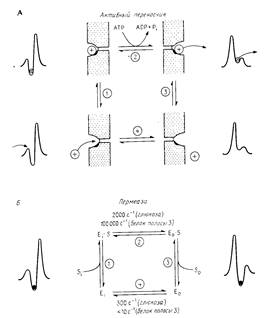
Рис.2
Рассмотрим четыре состояния белка-переносчика: