Курсовая работа: Водный режим у растений
Важное значение для развития корневых систем имеет аэрация . Именно недостаток кислорода является причиной плохого развития корневых систем на заболоченных почвах. Растения, приспособленные к росту на плохо аэрируемых почвах, имеют в корнях систему межклетников, которые вместе с межклетниками в стеблях и листьях составляют единую вентиляционную систему.
Большое значение имеют условия питания . Показано, что внесение фосфорных удобрений способствует углублению корневых систем, а внесение азотных удобрений – их усиленному ветвлению.
1.2 Основные двигатели водного тока
Поглощение воды корневой системой идет благодаря работе двух концевых двигателей водного тока: верхнего концевого двигателя, или присасывающей силы испарения (транспирации) , и нижнего концевого двигателя, или корневого двигателя. Основной силой, вызывающей поступление и передвижение воды в растении, является присасывающая сила транспирации, в результате которого возникает градиент водного потенциала. Водный потенциал – это мера энергии, используемой водой для передвижения. Водный потенциал и сосущая сила одинаковы по абсолютному значению, но противоположны по знаку. Чем меньше насыщенность водой данной системы, тем меньше (более отрицателен) ее водный потенциал. При потере воды растением в процессе транспирации создается ненасыщенность клеток листа водой, как следствие, возникает сосущая сила (водный потенциал падает) . поступление воды идет в сторону большей сосущей силы, или меньшего водного потенциала.[1]
Таким образом, верхний концевой двигатель водного тока в растении – это присасывающая сила транспирации листьев, и его работа мало связана с жизнедеятельностью корневой системы. Действительно, опыты показали, что вода может поступать в побеги и через мертвую корневую систему, причем в этом случае поглощение воды даже ускоряется.
Кроме верхнего концевого двигателя водного тока, в растениях существует нижний концевой двигатель. Это хорошо доказывается на примере таких явлениях, как гуттация.
Листья растений, клетки которых насыщены водой, в условиях высокой влажности воздуха, препятствующей испарению, выделяют капельно-жидкую воду с небольшим количеством растворенных веществ – гуттация. Выделение жидкости идет через специальные водные устьица – гидаторы. Выделяющаяся жидкость – гутта. Таким образом, процесс гуттации является результатом одностороннего тока воды, происходящего в отсутствие транспирации, и, следовательно, вызывается какой-то иной причиной.
К такому же выводу можно прийти и при рассмотрении явления плач растений. Если срезать побеги растения и к срезанному концу присоединить стеклянную трубку, то по ней будет подниматься жидкость. Анализ показывает, что это вода с растворенными веществами – пасока. В некоторых случаях, особенно в весенний период, плач наблюдается и при надрезе веток растений. Определения показали, что объем выделяющейся жидкости (пасоки) во много раз превышает объем корневой системы. Таким образом, плач – эт6о не просто вытекание жидкости в результате пореза. Все сказанное приводит к выводу, что плач, как и гуттация, связана с наличием одностороннего тока воды через корневые системы, не зависящего от транспирации. Силу, вызывающую односторонний ток воды по сосудам с растворенными веществами, не зависящую от процесса транспирации, называют корневым давлением. Наличие корневого давления позволяет говорить о нижнем концевом двигателе водного тока. Корневое давление можно измерить, присоединив манометр к концу, оставшемуся после срезания надземных органов растения, или поместив корневую систему в серию растворов различной концентрации и подобрав такую, при которой плач прекращается. Оказалось, что корневое давление равняется примерно 0,1 – 0,15 МПа (Д.А.Сабинин). Определения, проведенные советскими исследователями Л.В.Можаевой, В.Н.Жолкевичем, показали, что концентрация наружного раствора, останавливающего плач, значительно выше концентрации пасоки. Это позволило высказать мнение, что плач может идти против градиента концентрации. Было показано также, что плач осуществляется только в тех условиях, в которых нормально протекают все процессы жизнедеятельности клеток. Не только умерщвление клеток корня, но и снижение интенсивности их жизнедеятельности, в первую очередь интенсивность дыхания, прекращает плач. В отсутствии кислорода, под влиянием дыхательных ядов, при понижении температуры плач приостанавливается. Все сказанное позволило Д.А.Сабинину дать следующее определение: плач растений – это прижизненный односторонний ток воды и питательных веществ, зависящий от аэробной переработки ассимелятов. Д.А.Сабинин предложил схему, объясняющую механизм одностороннего тока воды в корне. Согласно этой гипотезе, клетки корня поляризованы в определенном направлении. Это проявляется в том, что в разных отсеках одной и той же клетки процессы обмена веществ различны. В одной части клетки идут усиленные процессы распада, в частности, крахмала на сахара, вследствие чего концентрация клеточного сока возрастает. На противоположном конце клетки преобладают процессы синтеза, благодаря чему концентрация растворенных веществ в этой части клетки уменьшается. Надо учитывать, что все эти механизмы будут работать только при достаточном количестве воды в среде и не нарушенном обмене веществ.[1]
Согласно другой гипотезе, зависимость плача растений от интенсивности дыхания является косвенной. Энергия дыхания используется для поступления ионов в клетки коры, откуда они десорбируются в сосуды ксилемы. В результате концентрация солей в сосудах ксилемы повышается, что и вызывает поступление воды.
1.3 Передвижение воды по растению
Вода, поглощенная клетками корня, под влиянием разности водных потенциалов, которые возникают благодаря транспирации, а также силе корневого давления, передвигается до проводящих путей ксилемы. Согласно современным представлениям, вода в корневой системе передвигается не только по живым клеткам. Еще в 1932 г. немецкий физиолог Мюнх развил представление о существовании в корневой системе двух относительно не зависимых друг от друга объемов, по которым передвигается вода, - апопласта и симпласта. Апопласт – это свободное пространство корня, в которое входят межклетные промежутки, оболочки клеток, а также сосуды ксилемы. Симпласт – это совокупность протопластов всех клеток, отграниченных полупроницаемой мембраной. Благодаря многочисленным плазмодесмам, соединяющим между собой протопласт отдельных клеток, симпласт представляет единую систему. Апопласт, по-видимому, не непрерывен, а разделен на два объема. Первая часть апопласта расположена в коре корня до клеток эндодермы, вторая – по другую сторону клеток эндодермы, и включает в себя сосуды ксилемы. Клетки эндодермы благодаря пояскам Каспари представляют как бы барьер для передвижения воды по свободному пространству (межклетникам и клеточным оболочкам). Для того чтоб попасть в сосуды ксилемы, вода должна пройти через полупроницаемую мембрану и главным образом по апопласту и лишь частично по симпласту. Однако в клетках эндодермы передвижение воды идет, по-видимому, по симпласту. Далее вода поступает в сосуды ксилемы. Затем передвижение воды идет по сосудистой системе корня, стебля и листа.[4]
Из сосудов стебля вода движется через черешок или листовое влагалище в лист. В листовой пластинке водопроводящие сосуды расположены в жилках. Жилки, постепенно разветвляясь, становятся более мелкими. Чем гуще сеть жилок, тем меньшее сопротивление встречает вода при передвижении к клеткам мезофилла листа. Иногда мелких ответвлений жилок листа так много, что они подводят воду почти к каждой клетке. Вся вода в клетке находится в равновесном состоянии. Иначе говоря, в смысле насыщенности водой, имеется равновесие между вакуолью, цитоплазмой и клеточной оболочкой, их водные потенциалы равны. Вода передвигается от клетки к клетке благодаря градиенту сосущей силы.
Вся вода в растении представляет единую взаимосвязанную систему. Поскольку между молекулами воды имеются силы сцепления (когезия), вода поднимается на высоту значительно большую 10 м. сила сцепления увеличивается, так как молекулы воды обладают большим сродством друг к другу. Силы сцепления обладают и между водой и стенками сосудов.[5]
Степень натяжения водных нитей в сосудах зависит от соотношения процессов поглощения и испарения воды. Все это позволяет растительному организму поддерживать единую водную систему и не обязательно восполнять каждую каплю испаряемой воды.
В том случае, если в отдельные членики сосудов попадает воздух, они, по-видимому, выключается из общего тока проведения воды. Таков путь передвижения воды по растению (рис. 2).
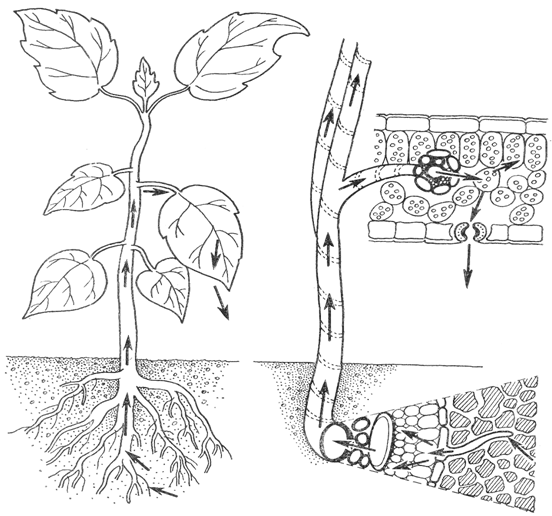
Рис.2. Путь воды в растении.
Скорость перемещения воды по растению в течение суток изменяется. В дневные часы она на много больше. При этом разные виды растений различаются по скорости передвижения воды. Изменение температуры, введение метаболических ингибиторов не влияют на передвижение воды. Вместе с тем этот процесс, как и следовало ожидать, очень сильно зависит от скорости транспирации и от диаметра водопроводящих сосудов. В более широких сосудах вода встречает меньшее сопротивление. Однако надо учитывать, что в более широкие сосуды могут попасть пузырьки воздуха или произойти какие-либо иные нарушения тока воды.[1]
2.Транспирация: физиологические механизмы
В основе расходования воды растительным организмом лежит физиологический процесс испарения – переход воды из жидкого в парообразное состояние, происходящей при соприкосновении органов растения с не насыщенной водой атмосферой. Однако этот процесс осложнен физиологическими и анатомическими особенностями растения, и его называют транспирацией
2.1 Назначение транспирации
В обычно протекающих процессах транспирация не является необходимой. Так если выращивать растения в условиях высокой и низкой влажности воздуха, то, естественно, в первом случае транспирация будет идти сознательно меньшей интенсивностью. Однако рост растений будет одинаков или даже лучше там, где влажность воздуха выше, а транспирация меньше. Известно, что большая часть всей поглощенной энергии тратится на транспирацию, которая в определенном объеме полезна растительному организму.[6]
1.Транспирация спасает растение от перегрева, который ему грозит на прямом солнечном свете. Температура сильно транспирирующего листа может быть примерно 7`C ниже температуры листа завязающего, не транспирирующего. Это особенно важно в связи с тем, что перегрев, разрушая хлоропласты, резко снижает процесс фотосинтеза (оптимальная температура для процесса фотосинтеза около 30-33`C). Именно благодаря высокой транспирирующей способности многие растения хорошо переносят повышенную температуру.
2.Транспирация создает непрерывный ток воды из корневой системы к листьям, который связывает все органы растения в единое целое.
3. С транспирационным током передвигаются растворимые минеральные и частично органические питательные вещества, при этом, чем интенсивнее транспирация, тем быстрее идет процесс передвижения.[6]
2.2 Лист как орган транспирации
Основным транспортирующим органом является лист. Средняя толщина листа составляет 100-200 мкм. Паренхимных клетки листа расположены рыхло, между ними имеется система межклетников, составляющая в общей сложности от 15 до 25% объема листа. Лист окружен покровной тканью- эпидермисом, состоящим из компактно расположенных клеток, наружные стенки которых утолщены. Листья большинства растений покрыта кутикулой, в состав которой входит оксимонокарбоновые кислоты, содержащие по 16-18 атомов углерода и по 2-3 гидроксильных группы. Эти кислоты соединены друг с другом в цепочки с помощью эфирных связей. Кутикула варьирует как по составу, так и по толщине. Более развитой кутикулой характеризуются листья светолюбивых растений по сравнению с влаголюбивыми. Кутикула вместе с клетками эпидермиса образует как бы барьер на пути испарения паров воды. При этом особенно значительную преграду составляет кутикула. Удаление кутикулы во много раз повышает интенсивность испарения. Сопротивление выходу паров оказывают в определенной мере и утолщенные стенки клетки эпидермиса. Все эти особенности выработались в процессе эволюции как приспособление к сокращению испарения. Для соприкосновения листа с атмосферой имеются устьица. Устьица – одно из оригинальных приспособлений листа, обладающее способностью открываться и закрываться. Обычно устьичные отверстия ограничены двумя замыкающими клетками, стенки которых не равномерно утолщены. У двудольных растений замыкающие клетки бобовидной, или полулунной, формы, при этом их внутренние прилегающие друг к другу стенки долее толстые, а внешние – более тонкие. Когда воды мало, замыкающие клетки плотно прилегают друг к другу и устьичная щель закрыта. Когда воды в замыкающих клетках много, то она давит на стенки и более тонкие стенки растягиваются сильнее, а более толстые втягиваются внутрь, между замыкающими клетками появляется щель. У однодольных растений строение замыкающих клеток несколько иное. Они представлены двумя удлинёнными клетками, на концах которых стенки более тонкие. При насыщении водой более тонкие стенки на концах растягиваются и раздвигают замыкающие клетки, благодаря чему образуется щель. (рис.3)
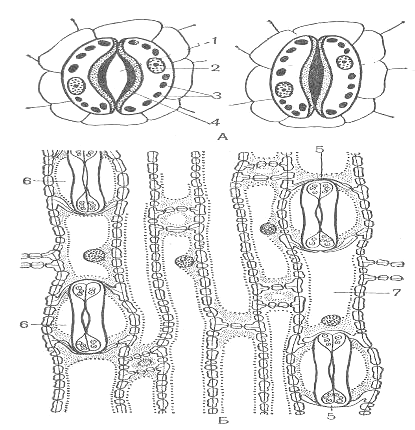
Рис.3 Структура устьиц у двудольных (А) и однодольных (Б) растений: