Реферат: Филогенез и онтогенез коры больших полушарий
На рисунке 1 представлена схема цитоархитектонического строения новой коры. В разделе А окрашены только тела клеток, Б окрашены отдельные клетки с отростками; В окрашены волокна. Слои коры: I — молекулярный слой; II — наружный зернистый слой; III — пирамидный слой; IV — внутренний зернистый слой; V — ганглионарный слой; VI — полиморфный слой.
Новая кора
Все области новой коры построены по единому принципу. Исходным типом является шестислойная кора (рис. 1). Слои представлены следующим образом:
Iслой — самый поверхностный, толщиной около 0,2 мм, называется молекулярным (lamina molecula ris). Он состоит из волокон апикальных дендритов и аксонов, поднимающихся от клеток нижних слоев, которые контактируют друг с другом. Нейронов в молекулярном слое незначительное количество. Это мелкие горизонтальные клетки и клетки-зерна. Все отростки клеток молекулярного слоя располагаются в пределах этого же слоя.
IIслой— наружныйзернистый(lamina granula ns exte rna).Толщина наружного зернистого слоя — 0,10 мм. Он состоит из мелких пирамидных и звездчатых нейронов. Аксоны этих нейронов оканчиваются на нейронах III, V и VI слоев.
III слой — пирамидный {lamina pyramidalis),толщиной около 1 мм, состоит из мелких и средних пирамидных клеток. Типичный пирамидный нейрон имеет форму треугольника, вершина которого направлена вверх. От вершины отходит апикальный дендрит, ветвящийся в вышележащих слоях. Аксон пирамидной клетки отходит от основания клетки и направляется вниз. Дендриты клеток III слоя направляются во второй слой. Аксоны клеток III слоя оканчиваются на клетках нижележащих слоев или образуют ассоциативные волокна.
IV слой — внутренний зернистый {lamina granula ns inte rnus).Он состоит из звездчатых клеток, имеющих короткие отростки, и малых пирамид. Дендриты клеток IV слоя уходят в молекулярный слой коры, а их коллатерали ветвятся в своем слое. Аксоны клеток IV слоя могут подниматься в вышележащие слои или уходить в белое вещество как ассоциативные волокна. Толщина IV слоя от 0,12 до 0,3 мм.
V слой — ганглионарный {lamina gangliona ris) — слой больших пирамид. Самые крупные клетки коры расположены именно в этом слое (гигантские пирамиды Беца передней центральной извилины). Их апикальные дендриты достигают молекулярного слоя, а базальные дендриты распределяются в своем слое. Аксоны клеток V слоя покидают кору и являются ассоциативными, комиссуральными или проекционными волокнами. Толщина V слоя достигает 0,5 мм.
VI слой коры — полиморфный {lamina multifo rmis).Содержит клетки разнообразной формы и размера, имеет толщину от 0,1 до 0,9 мм. Часть дендритов клеток этого слоя достигает молекулярного слоя, другие же остаются в пределах IV и V слоев. Аксоны клеток VI слоя могут подниматься к верхним слоям или уходить из коры в качестве коротких или длинных ассоциативных волокон.
Клетки одного слоя коры выполняют сходную функцию в обработке информации. I и IV слои являются местом ветвления ассоциативных и комиссуральных волокон, т.е. получают информацию от других корковых структур. III и IV слои являются входными, афферентными для проекционных полей, так как именно в этих слоях заканчиваются таламические волокна. V слой клеток выполняет эфферентную функцию, его аксоны несут информацию к нижележащим структурам мозга. VI слой также является выходным, но его аксоны кору не покидают, а являются ассоциативными.
Основным принципом функциональной организации коры является объединение нейронов в колонки. Колонка расположена перпендикулярно поверхности коры и охватывает все ее слои от поверхности к белому веществу. Связи между клетками одной колонки осуществляются по вертикали вдоль оси колонки. Боковые отростки клеток имеют небольшую длину. Связь между колонками соседних зон осуществляется через волокна, уходящие вглубь, а затем входящие в другую зону, т.е. короткие ассоциативные волокна. Функциональная организация коры в виде колонок обнаружена в соматосенсорной, зрительной, моторной и ассоциативной коре. Отдельные зоны коры имеют принципиально одинаковое клеточное строение, однако есть и отличия, особенно в структуре III, IV и V слоев, которые могут распадаться на несколько подслоев. Кроме этого, существенными цитоархитектоническими признаками являются плотность расположения и размеры клеток, наличие специфических типов нейронов, расположение и направление хода миелиновых волокон.
Цитоархитектонические особенности позволили разделить всю поверхность коры на 11 цитоархитектонических областей, включающих в себя 52 поля (по Бродману). Каждое цитоархитектоническое поле обозначено на картах мозга номером, который присваивался ему в порядке описания.
В первом слое коры волокна образуют полоску молекулярной пластинки. Во втором слое залегают тонкие волокна наружной зернистой пластинки. В составе четвертого слоя коры находится полоска внутренней зернистой пластинки (наружная полоска Байярже). В пятом слое содержатся волокна внутренней пирамидной пластинки (внутреняя полоска Байярже).
Основная информация в кору поступает по специфическим афферентным проводящим путям, заканчивающимся на клетках 3 и 4 слоев. Неспецифические пути от РФ заканчиваются в верхних слоях коры и регулируют ее функциональное состояние (возбуждение, торможение).
Звездчатые нейроны выполняют главным образом чувствительную (афферентную) функцию. Пирамидные и веретеновидные клетки – это преимущественно двигательные (эфферентные) нейроны.
Часть клеток коры принимают информацию от любых рецепторов организма – это полисенсорные нейроны, воспринимающие импульсы только от определенных рецепторов (зрительных, слуховых, тактильных и т.д.). Клетки нейроглии выполняют вспомогательные функции: трофическую, нейросекреторную, защитную, изолирующую.
Специализированные нейроны и другие клетки, входящие в состав вертикальных колонок, образуют отдельные участки коры, которые называются проекционными зонами (цитоархитектоническими полями)[1] . Эти функциональные зоны коры делятся на 3 группы:
– афферентные (чувственные);
– эфферентные (двигательные или моторные);
– ассоциативные (соединяют предыдущие зоны и обусловливают сложную работу мозга, лежащую в основе высшей психической деятельности).
Ассоциативные зоны и локализация полей
Каждый анализатор (например, зрения, обоняния, слуха и т.д.) имеет, по представлению И. П. Павлова, в коре головного мозга центральную часть (ядро), где осуществляется высший анализ и синтез, и широкую периферическую зону, в которой аналитические и синтетические процессы совершаются в элементарном виде. Между ядрами отдельных анализаторов разбросаны и перемешаны нервные элементы, принадлежащие различным анализаторам. Если ядро анализатора в силу каких-либо причин разрушено или выбыло из строя, его функцию перенимают периферические элементы того же анализатора. Современная физиология отвергает и узкий («абсолютный») локализационизм и принцип однородности, равноценности всех участков коры мозга. Локализация существует, но имеет «подвижный», «динамический» характер, о чем еще много лет назад говорил И.П. Павлов.
Нервные образования, которые мы привыкли называть «центрами», не ограничиваются корой головного мозга. Они включают и подкорковые структуры, значение которых необычайно велико.
Следует помнить, что любой центр коры головного мозга теснейшим образом связан со всеми другими отделами центральной нервной системы. В этом объединении, или, как говорят, интеграции, и заключается ведущая роль коры мозга в организме. Представление о единых корковых центрах, полностью обеспечивающих какую-либо определенную функцию, является в настоящее время пройденным этапом в физиологии.
К тому же кора головного мозга отличается необычайной пластичностью, и одни отделы ее легко перенимают функции других, компенсируя расстройство их деятельности, вызванное различными причинами. Наиболее важная задача современной науки выявить анатомическую основу физиологических процессов и одновременно установить связи и взаимосвязи между всеми явлениями, наблюдаемыми в головном мозгу. Исследования, проведенные различными авторами, как отечественными, так и зарубежными, показали, что в центральной извилине мозга, расположенной спереди от центральной борозды, находится специальная двигательная область. Раздражение ее электрическим током вызывает сокращение определенных мышц противоположной стороны тела. Напротив, удаление этой области хирургическим путем ведет к расстройству координированных движений, шаткости походки, ослаблению мышц. У человека ранение двигательной области сопровождается обычно параличами и другими тяжелыми нарушениями деятельности организма. С помощью метода условных рефлексов удалось показать, что так называемые двигательные центры содержат чувствительные клетки, к которым приходят периферические раздражения от двигательного аппарата (костей, суставов, мышц). Эта область является мозговым концом двигательного анализатора в такой же степени, как затылочная — мозговым концом зрительного анализатора, височная — слухового анализатора и т.д. В двигательной области имеются как чувствительные клетки, расположенные в верхних слоях коры, так и двигательные, сосредоточенные в ее нижних слоях. Импульсы от рецепторов двигательного аппарата поступают в чувствительные клетки передней мозговой извилины, а отсюда уже передаются двигательным клеткам головного и спинного мозга.
У человека ассоциативные зоны достигают наибольшего развития. Локализация функций в коре головного мозга относительна – здесь нельзя провести каких-либо четких границ, поэтому мозг обладает высокой пластичностью, приспосабливаемостью к повреждениям. Тем не менее, морфологическая и функциональная неоднородность коры позволила выделить в ней 52 цитоархитектонических поля (К. Бродман), а среди них – центры зрения, слуха, осязания и др. Все они связаны между собой волокнами проводящих путей белого вещества, которые делятся на 3 типа:
1. ассоциативные (связывают зоны коры в пределах одного полушария),
2. комиссуральные (связывают симметричные зоны коры двух полушарий через мозолистое тело),
3. проекционные (связывают кору и подкорку с периферическими органами, бывают чувствительные и двигательные).
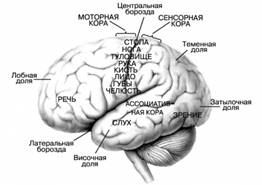
Рисунок 2. Значение важнейших зон коры головного мозга