Реферат: Гистогенез, морфо-функциональные и гисто-химические особенности мышечной ткани. Механизм мышечного сокращения
Миоциты этой ткани развиваются из клеток нейрального зачатка в составе внутренней стенки глазного бокала. Тела этих клеток располагаются в эпителии задней поверхности радужки. Каждая из них имеет отросток, который направляется в толщу радужки и ложится параллельно ее поверхности. В отростке находится сократительный аппарат, организованный так же, как и во всех гладких миоцитах. В зависимости от направления отростков (перпендикулярно или параллельно краю зрачка) миоциты образуют две мышцы – суживающую и расширяющую зрачок.
Сокращение мышц
Теория скольжения нитей
Н.Е. Huxley и A.F. Huxley независимо друг от другав 1954 г. предложили для объяснения механизма мышечного сокращения теорию скольжения нитей. Согласно данной теории, укорочение саркомера, а, следовательно, и мышечного волокна в момент сокращения происходит благодаря активному скольжению тонких (актиновых) нитей относительно толстых (миозиновых) нитей. Укорочение заканчивается, когда актиновые филаменты глубоко втягиваются по направлению к центру диска, который определяет границы саркомеров. При расслаблении или растяжении мышцы область взаимного перекрывания тонких и толстых филаментов сужается.
Скользящее движение миозиновых и актиновых филаментов друг относительно друга обусловлено силами, генерируемыми при взаимодействии поперечных мостиков с актиновыми филаментами.
Поперечные мостики должны последовательно прикрепиться к актиновому филаменту, развить силу, отойти и вновь прикрепиться в другом месте. Для того чтобы поддерживать активное сокращение, поперечные мостики должны работать асинхронно, т.е. в любой момент времени часть из них прикреплена к актину, тогда как другие отсоединены. После отсоединения поперечный мостик должен вновь прикрепиться к актиновому филаменту, но уже дальше, в сторону Z-пластинок, внося тем самым вклад в активное скольжение вдоль указанного направления.
Один из основных вопросов по поводу функционирования поперечных мостиков относится к преобразованию химической энергии в механическую. Как же все-таки поперечные мостики генерируют силу для скольжения толстых и тонких филаментов друг относительно друга? По этому поводу высказан ряд гипотез. Широкое распространение получила точка зрения, что сила генерируется за счет колебания или вращения миозиновой головки и затем передается на толстую нить через шейку молекулы миозина. Шейка образует мостиковый шарнир, расположенный между головкой миозиновой молекулы и толстым филаментом. В данной гипотезе мостиковый шарнир выступает как соединение между головкой миозина и толстым филаментом, которое передает силу, развиваемую при вращении головки на актиновом филаменте.
Исследования механических свойств сокращающейся мышцы, проведенные Хаксли и Симмонсом, подтвердили такую точку зрения на функцию поперечных мостиков. Авторы показали, что основная часть упругого компонента мышцы, включенная последовательно с сократительным элементом, находится в самих поперечных мостиках, предположительно в мостиковом шарнире. Они высказали мысль, что упругое растяжение шарнира служит важным моментом в процессе запасания механической энергии при вращении головки миозина вокруг актинового филамента. В соответствии с данной гипотезой вращение генерируется несколькими центрами миозиновой головки, которые поочередно взаимодействуют с центрами на актиновом филаменте.
Упругость мостикового шарнира способствует вращению головки без заметных скачкообразных колебаний развиваемой силы. Растянувшись, мостиковый шарнир будет передавать свое усилие толстому филаменту мягко, содействуя активации скольжения филаментов. Один из главных аргументов-это то, что, по данным Хаксли и Симмонса, последовательно соединенный упругий компонент мышечного волокна пропорционален величине взаимного перекрывания тонких и толстых филаментов, а следовательно, пропорционален числу присоединенных поперечных мостиков. Авторы также установили, что внезапно возникающее небольшое укорочение сопровождается очень быстрым возрастанием развиваемого усилия; они объясняют это лишь поворотом головок поперечных мостиков, взаимодействующих с актином, в более стабильное положение.
Роль кальция в процессе сокращения
Данные о роли ионов кальция в сократительной активности мышц накапливались довольно медленно. Кальций активен в саркоплазме при такой низкой (10-6 М и менее) концентрации, что до открытия кальцийхелатных реагентов, например ЭДТА и ЭГТА, ее невозможно было поддерживать в экспериментальных растворах. Дело в том, что даже в бидистиллированной воде концентрация ионов кальция превышает 10-6 М. Самые первые доказательства физиологической роли Са2+ представлены в работах Рингера и Бакстона. Авторы обнаружили, что изолированное сердце лягушки прекращает сокращения при отсутствии кальция в омывающем растворе. Так появились раствор Рингера и другие физиологические солевые растворы.
Камада и Киносита, а затем Хейлбрун и Вертинский проверяли участие Са2+ в регуляции мышечного сокращения путем введения разных катионов внутрь мышечных волокон. Из всех изученных ионов только кальций вызывал сокращение при концентрациях, соизмеримых с концентрациями Са2+ обычно наблюдаемыми в живой ткани. Впоследствии было обнаружено, что скелетная мышца не сокращается в ответ на деполяризацию мембраны, если исчерпаны запасы кальция во внутренних депо, а подвергнутые предварительной экстракции препараты волокон скелетной мышцы не сокращаются при добавлении АТФ, если отсутствует Са2+ .
Количественная зависимость между концентрацией свободного Са2+ в саркоплазме и силой мышечного сокращения была установлена сравнительно недавно. Для проведения анализа удаляли поверхностную мембрану и оголенные миофибриллы обрабатывали растворами кальция различной концентрации. Сила возрастает от нуля при концентрации кальция около 10-8 М до максимального значения при концентрации кальция около 5х10-6 М. Данная зависимость между силой и концентрацией Са2+ аналогична зависимости между АТФазной активностью (скоростью гидролиза АТФ) гомогенизированных миофибрилл и концентрацией Са2+ . Такое совпадение характеристик наводило на мысль, что Са2+ служит кофактором АТФазной активности миозина. Но оказалось, что это не так.
АТФазная активность чистого раствора миозина довольно низкая, но сильно возрастает при добавлении очищенного актина. Это указывает на то, что АТФазный центр миозина активируется при связывании миозина с актином. В интактной мышце активация АТФазного центра миозина осуществляется при присоединении поперечного мостика к активному филаменту. Эксперименты, проведенные в лаборатории Эбаши, показали, что тропонин и тропомиозин, лежащие вдоль актиновой спирали, препятствуют присоединению миозиновых поперечных мостиков к актину. Тропонин – единственный белок в актиновых и миозиновых филаментах поперечнополосатых мышц позвоночных животных, имеющий высокое химическое сродство к Са2+ . Каждый тропониновый комплекс связывает четыре иона кальция. Тропониновые комплексы расположены вдоль актинового филамента через каждые 40 нм, прикрепляясь одновременно к актиновому филаменту и молекуле тропомиозина. В состоянии покоя положение тропомиозина конформационно препятствует соединению головок миозина с актиновым филаментом. Связывая Са2+ , тропонин претерпевает конформационные изменения, в результате чего молекула тропомиозина смещается и освобождает дорогу миозиновым поперечным мостикам для прикрепления к актиновым центрам. Следовательно, присоединение Са2+ к тропонину устраняет постоянно существующее препятствие для взаимодействия поперечных мостиков с актином. Из результатов экспериментов, сделан вывод, что ингибирование присоединения мостиков снимается при концентрации свободного Са2+ свыше 10-7 М.
Сказанное выше объясняет роль Са2+ в регуляции актин-миозинового взаимодействия в скелетных и сердечной мышце позвоночных животных. В большинстве других мышц роль кальция иная. Есть еще по крайней мере два механизма кальцийзависимой регуляции актин-миозинового взаимодействия. В поперечнополосатых мышцах большинства беспозвоночных животных кальций инициирует сокращение, присоединяясь к легким полипептидным цепям миозина в головках поперечных мостиков. В гладких мышцах позвоночных животных и в немышечном актомиозине сокращение контролируется кальцийзависимым фосфорилированием миозиновой головки.
Инактивация поперечных мостиков и расслабление мышцы
В мышце, находящейся в состоянии покоя, внутренняя система ограниченных мембранами компартментов, называемая саркоплазматическим ретикулумом , активно поглощает Са2+ . Благодаря этому процессу уровень свободных ионов кальция не поднимается выше 10-7 М. При такой концентрации поперечные мостики неактивны, потому что с тропонином связывается лишь очень небольшое количество кальция. Таким образом, удаление Са2+ из саркоплазмы в ретикулуме заставляет мышцу расслабляться после сокращения.
Поскольку АТФ поставляет энергию для сокращения, напрашивается вывод, что удаление АТФ тоже вызовет расслабление мышцы. Но оказалось, что этого не происходит.
Мышца становится напряженной и не поддается растяжению при исчерпании всех ее запасов АТФ и фосфагенов. Это состояние известно как трупное окоченение , и обусловлено оно тем, что поперечные мостики не могут отделиться от актиновых филаментов. О том, что для расслабления мышцы нужен Мg2 + -АТФ, известно со времени проведения первых экспериментов с экстрагированными глицерином препаратами мышц. В присутствии Са2+ и Мg2 + -АТФ глицеринизированная мышца сокращается, а при удалении Са2+ – расслабляется. Расслабление, как и сокращение, происходит только в присутствии Мg2 + -АТФ. В нормальных условиях, когда мышца обеспечена АТФ, мостики легко отделяются. Затем, если концентрация свободного саркоплазматического Са2+ становится ниже уровня, необходимого для процесса присоединения поперечных мостиков к актиновым филаментам, мышца расслабляется.
Итак, расслабление мышцы зависит от наличия Мg2 + -АТФ, необходимого для разрушения актомиозинового комплекса, и от внутриклеточной концентрации кальция, которая должна быть достаточно низкой для предотвращения нового прикрепления мостиков к актиновым филаментам.
Саркоплазматический ретикулум
С чего начинается поступление Са2+ в СР? Если мембраны СР выделить с помощью фракционирования, они образуют микроскопические везикулы диаметром 1 мкм. Везикулы способны поглощать кальций из окружающей среды. Если к ним добавить щавелевую кислоту, то внутри везикул по мере увеличения в них концентрации Са2+ будет осаждаться оксалат кальция. Это говорит об активном транспорте кальция мембраной ретикулума. В нефракционированной мышечной ткани осадок оксалата кальция можно обнаружить с помощью электронного микроскопа в терминальных цистернах. Способность СР к накоплению кальция довольно высокая, что обеспечивает поддержание концентрации свободного Са2+ в саркоплазме расслабленной мышцы ниже 10-7 М. Этот уровень Са2+ достаточен для разрушения связи кальция с тропонином и предотвращения сокращения. Способность СР поглощать Са2+ из миоплазмы зависит от активности молекул кальциевого насоса. На электронных микрофотографиях, полученных методом замораживания-скалывания, молекулы насоса плотно прижаты («плечом к плечу») в мембранах, формирующих продольные элементы СР. Как и в других активных транспортных системах, в качестве источника энергии кальциевый насос СР использует АТФ.
Высвобождение кальция саркоплазматическим ретикулумом
Как только стало известно, что в СР накапливаются ионы кальция, исследователи начали склоняться к мысли о том, что мышечное сокращение инициируется Са2+ , высвобождаемым в саркоплазму из внутренней среды цистерн СР.
Сокращение активируется кальцием, высвобожденным из СР, а поверхностный электрический сигнал, т.е. ПД, поступает в глубокие области мышечного волокна с помощью Т-трубочек. Более того, Т-трубочки образуют тесные контакты с концевыми цистернами саркоплазматического ретикулума. Но как электрический сигнал из Т-трубочек передается в СР, давая команду к высвобождению Са2+ в ответ на деполяризацию Т-трубочки, долгое время оставалось загадкой. Сейчас, кажется, на этот важный вопрос можно ответить. Очевидно, что при деполяризации Т-трубочек сигнал доставляется к концевым цистернам СР посредством внутриклеточных молекул-посредников. Недавние исследования, проведенные в Калифорнийском университете, показали, что высвобождение Са2+ из СР и последующее сокращение одиночного поперечного волокна могут индуцироваться инозитол-1,4,5- трифосфатом (ИФ3 ). Это внутриклеточная молекула-посредник, образующаяся при разложении связанного с мембраной фосфатидилинозитола, которая, как известно, стимулирует высвобождение Са2+ из внутриклеточных хранилищ в некоторых тканях. В отношении мышц есть сведения, что вещества, блокирующие образование ИФ3 , нарушают сопряжение процессов сокращения волокна и деполяризации мембран. Показано, что такими вещества мешают нормальному высвобождению Са2+ из СР в ответ на электрическое возбуждение мышцы. И наконец, вещества, блокирующие ферментативное разложение ИФ3 , напротив, усиливают эффективность ИФ3 , в инициации сокращения мышечного волокна. Такого рода данные послужили поводом для возникновения гипотезы, утверждающей, что деполяризация Т-трубочек вызывает образование ИФ3 , а уже затем ИФ3 , действует как внутриклеточный посредник, индуцирующий
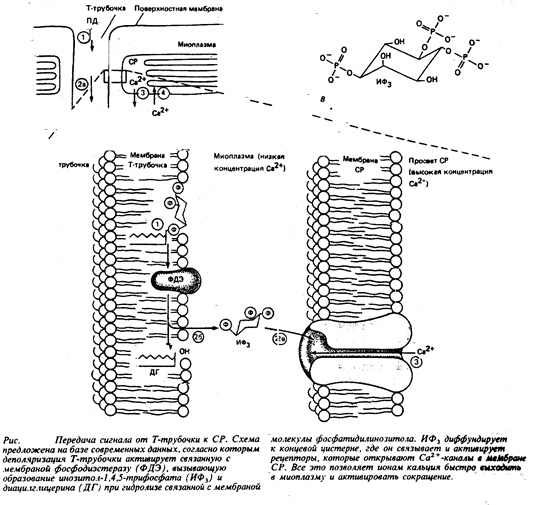 |
????????????? ??2+ ?? ?? (???.5).
Согласно этой гипотезе, начальная стадия сопряжения процесса «возбуждение – сокращение» сопровождается распространением возбуждения по поверхности системы Т-трубочек и представляет собой активацию чувствительных к электрическому напряжению ферментов, расположенных на мембране данных трубочек рядом с концевыми цистернами СР. Эти гипотетические ферменты, по-видимому, столь же чувствительны к изменению электрического поля мембраны, как натриевый канал, и реагируют на это изменение конформационным сдвигом. Вызванный деполяризацией мембраны конформационный сдвиг переводит фермент из неактивной формы в активную. И уже этот активный фермент прямо или косвенно определяет образование ИФ3 . Затем ИФ3 диффундирует на короткое расстояние и достигает мембраны концевой цистерны СР, где, связавшись с рецептором, заставляет открываться кальциевые каналы. Ионы кальция, скопившиеся в относительно высокой концентрации в просвете СР, продолжают выходить наружу до тех пор, пока не произойдет ферментативное разрушение ИФ3 и каналы не закроются. Потом с помощью активного транспорта высвобожденные из СР ионы кальция возвращаются на прежнее место.
Краткое описание процессов сокращения и расслабления
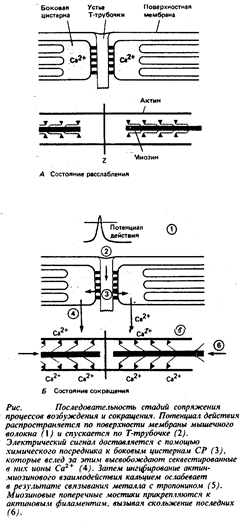 Процессы, контролирующие сокращение скелетной мышцы, изображены в общем виде на рис.6. Приведем их перечень.
Процессы, контролирующие сокращение скелетной мышцы, изображены в общем виде на рис.6. Приведем их перечень.
1. Поверхностная мембрана мышечного волокна деполяризуется под влиянием потенциала действия или (в некоторых мышцах) под влиянием синаптических потенциалов.
2. Потенциал действия поступает в глубь мышечного волокна по Т-трубочкам.
3. В ответ на деполяризацию Т-трубочек сигнал, который, вероятно, опосредуется молекулами ИФ3, распространяется от этих трубочек к концевым цистернам саркоплазматического ретикулума.
4. Этот химический посредник вызывает открытие кальциевых каналов в СР и высвобождение секвестированных там ионов кальция.
5. Концентрация свободного Са2+ в миоплазме возрастает от значения 10-7 М и ниже (в покое) до приблизительно 10-6 М и более (в активном состоянии). Кальций соединяется с тропонином, вызывая в молекуле этого белка конформационные изменения.
6. Конформационные изменения молекулы тропомиозина устраняют пространственное препятствие для присоединения поперечных мостиков к актиновым филаментам.
7. Миозиновые поперечные мостики прикрепляются к актиновым филаментам и вступают в последовательное взаимодействие с их центрами, что вызывает вращение миозиновой головки относительно актиновых филаментов и натяжение мостикового шарнира.
8. Натяжение мостикового шарнира приводит к активному вхождению актиновых филаментов в А-диск. Саркомер слегка укорачивается.