Реферат: Костистые рыбы
Рис. 5 Череп судака сбоку: 1 — окологлазничные кости, 2— слезная кость, 3 — нёбная кость, 4 — квадратная кость, 5 — крыловидные кости, 6 — сочленовная кость, 7 — предчелюстная кость, 8 — верхнечелюстная кость, 9 — зубная кость, 10 — угловая кость, 11 — подвесок, 12 — симплектикум, 13 — гиоид, 14 — копулагиоида, 15 — крышечная кость, 16 — предкрышечная кость, 17 — подкрышечная кость, 18 — межкрышечная кость
При рассматривании черепа сбоку помимо уже отмеченных костей видно полукольцо из мелких косточек, окаймляющих глазницу — окологлазничных (ossa infraorbitalia; рис. 5, 1). Передняя из них носит особое название — слезная кость (lacrimale; рис. 5, 2). Все эти кости покровные.
Висцеральный череп костистых рыб представлен серией висцеральных дуг: челюстной (самая передняя), подъязычной и пятью жаберными, из которых последняя (задняя) заметно редуцирована.
В челюстной дуге можно различить первичные и вторичные челюсти. Первичные челюсти образованы преимущественно замещающими костями и гомологичны хрящам нёбно-квадратному и меккелеву хрящевых рыб. В верхней челюсти они представлены спереди смешанной нёбной костью (palatinum; рис. 5, 3), сзади (в месте сочленения с нижней челюстью) — квадратной костью (quadratum; рис. 5, 4); между ними расположены три крыловидные кости (pterygoideum; рис. 33, 5). Из них лишь одна (задняя крыловидная — metapterygoicieum) замещающая, тогда как остальные две — наружная крыловидная (ectopterygoideum) и внутренняя крыловидная (entopterygoideum) — по происхождению покровные.
В нижней челюсти первична лишь одна сочленовная кость (articulare; рис. 5, 6), осуществляющая подвижное причленение нижней челюсти к верхней (через квадратную кость).
Вторичные челюсти представлены предчелюстными (praemaxillare; рис. 5, 7) и верхнечелюстными (maxillare; рис. 5, 8) костями в верхней половине и крупной зубной костью (dentale, рис. 5, 9) — в нижней. Кроме того, в нижней челюсти имеется еще маленькая угловая кость (angulare; рис. 5, 10), приросшая к задне-нижнему краю сочленовной. Предчелюстная и зубная кости снабжены довольно крупными зубами. Вторичным челюстям принадлежит главная хватательная функция. Дополнительную функцию удерживания добычи выполняют зубы, расположенные на нёбных костях и сошнике.
Все перечисленные кости челюстной дуги — парные; они симметрично повторяются на правой и левой половинах челюстей.
Подъязычная дуга целиком составлена первичными (замещающими) костями. Верхний элемент ее — подвесок (hyomandibulare, рис. 5, 11) верхним концом причленяется к слуховому отделу осевого черепа, а нижним — через маленькую дополнительную косточку (symplecticum; рис. 5, 12) — к квадратной кости челюстной дуги. Кроме этого, подвесок срастается с задней крыловидной костью. Таким образом, подвесок выполняет функцию причленения челюстной дуги к осевому черепу; иными словами, череп у костистых рыб, как и у акуловых, гиостилический. Нижняя ветвь подъязычной дуги, гомологичная гиоидному хрящу акуловых рыб, представлена несколькими окостенениями. Из них крупный — гиоид (hyoideum; рис. 5, 13). Гиоиды правой и левой сторон соединяются через непарную кость (copula; рис. 5, 14), одновременно поддерживающую язык. К гиоиду прикрепляются тонкие изогнутые косточки — лучи жаберной перепонки, поддерживающие кожистый край жаберной крышки.
Жаберные дуги представлены каждая четырьмя парными косточками, подвижно сочлененными между собой и объединяющимися снизу при помощи непарных костных элементов — копул. Исключение составляет лишь пятая, самая задняя жаберная дуга, имеющая лишь один парный (нижний) отдел.
Жаберные крышки являются по сравнению с хрящевыми рыбами новообразованием. Каждая состоит из четырех вторичных (покровных) костей: крышки (operculum; рис. 5, 15), предкрышки (praeoperculum; рис. 5, 16), подкрышки (suboperculum; рис. 5, 17) и межкрышки (interoperculum; рис. 5, 18). Жаберная крышка каждой стороны через предкрышечную кость прикрепляется к соответствующему подвеску и к квадратной кости.
Парные конечности и их пояса. Парные конечности представлены грудными и брюшными плавниками. Опорой грудных плавников в теле рыбы служит плечевой пояс (рис. 6). Он представлен двумя небольшими замещающими (первичными) и несколькими покровными костями. Верхняя из замещающих костей — лопатка (scapula; рис. 6, 1) — расположена в области причленения свободной конечности (ее легко отличить по небольшому круглому отверстию в центре кости). Сразу под ней находится врановая кость, или коракоид (coracoideum; рис. 6, 2). Эти два элемента составляют первичный пояс. Они неподвижно соединены с крупной покровной костью клейтрум (cleithrum; рис. 6, 3), верхний конец которой направлен несколько вперед; к нему присоединяется небольшая кость надклейтрум (supracleithrum; рис. 6, 4).
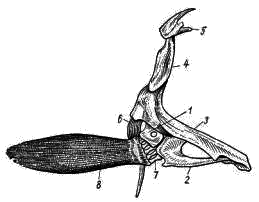
Рис. 6. Плечевой пояс и грудной плавник судака: 1 — лопатка, 2 — коракоид, 3 — клейтрум, 4 — надклейтрум, 5 — заднетеменная кость, 6 — заднеключичная кость, 7 — радиалии, 8 — костные кожные лучи
Клейтрум в свою очередь соединяется с заднетеменной (posttemporale; рис. 6, 5) костью. Направленные вперед нижние концы правого и левого клейтрумов соединяются друг с другом. Позади клейтрума неподалеку от лопатки и коракоида расположена небольшая заднеключичная кость (postcleithrum; рис. 6, 6). Все названные кости парные; они составляют вторичный плечевой пояс. Правая и левая заднетеменные кости причленяются к осевому черепу, что обеспечивает более прочную фиксацию пояса и таким образом усиливает его опорную функцию.
Грудной плавник в своем основании имеет один ряд мелких косточек — радиалии (radialia; рис. 6, 7), отходящих от лопатки (частично и от коракоида). Вся свободная лопасть плавника состоит из членистых кожных лучей1 (lepidotrichia; рис. 6, 8). Особенность скелета грудных плавников костистых рыб, по сравнению с хрящевыми, заключается в редукции базалий. Подвижность грудных плавников увеличивается потому, что мышцы прикрепляются к расширенным основаниям кожных лучей, подвижно сочленяющихся с радиалями.
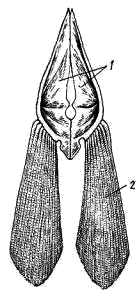
Рис. 7. Тазовый пояс и брюшной плавник судака: 1 — тазовые кости, 2 — кожные костные лучи
Тазовый пояс (рис. 7) представлен сливающимися друг с другом парными плоскими треугольными костями, лежащими в толще брюшной мускулатуры и не связанными с осевым скелетом. К боковым сторонам тазового пояса причленяются брюшные плавники. У большинства костистых рыб в скелете брюшных плавников отсутствуют базалий и полностью редуцировались радиалии: лопасть плавника поддерживается кожными костными лучами (lepidotrichia), расширенные основания которых непосредственно причленяются к тазовому поясу. Такое упрощение скелета брюшных плавников, видимо, связано с их ограниченными функциями.
Непарные конечности. Непарные конечности представлены спинными, подхвостовым (анальным) и хвостовым плавниками. Анальный и спинные плавники состоят из костных лучей, подразделяющихся на внутренние (скрытые в толще мускулатуры) птеригофоры и наружные плавниковые лучи — лепидотрихии.
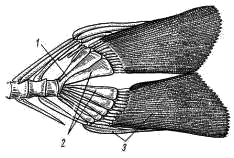
Рис. 8. Хвостовой плавник судака: 1 — конец позвоночного столба — уростиль, 2 — гипуралин, 3 — костные кожные лучи — лепидотрихии
Хвостовой плавник, как это было видно уже при наружном осмотре, имеет внешне равнолопастное строение, однако при рассмотрении его внутреннего скелета (рис. 8) видно, что концевые позвонки позвоночного столба сливаются в палочковидную косточку — уростиль (urostyl; рис. 8, 1), которая заходит в основание лишь верхней лопасти плавника, а основание нижней лопасти поддерживается разросшимися, довольно широкими нижними дугами позвонков — гипуралиями (рис. 8, 2). Такой тип строения хвостового плавника, вполне или почти симметричного по внешней форме, но асимметричного по расположению внутреннего скелета, носит название гомоцеркального. Наружный скелет хвостового плавника составлен многочисленными кожными лучами — лепидотрихиями (рис. 8, 3).
ЗАКЛЮЧЕНИЕ
Общий тип организации водных челюстноротых позвоночных, сформировавшийся у хрящевых рыб, претерпевает дальнейшее развитие в классе костных рыб. Сохраняя общий план строения предков, костные рыбы в своей эволюции приобрели ряд прогрессивных черт, способствовавших повышению уровня жизнедеятельности и широкой адаптивной радиации этих животных.
Наиболее существенное отличие костных рыб от хрящевых заключается в развитии у них костного скелета. Биологическое значение этого явления очень велико: превосходя хрящ в прочности, костная ткань придает скелету еще и большую легкость и подвижность.
Развитие костной ткани происходит двумя независимыми путями. С одной стороны, имеют место довольно сложные гистологические процессы, в результате которых хрящевая ткань, составляющая основу скелета зародыша, постепенно замещается костной. Так возникают замещающие (хрящевые, или первичные) кости. Первичный костный скелет принимает на себя практически все те функции, которые у хрящевых рыб принадлежали хрящевому скелету.
С другой стороны, костная ткань может формироваться непосредственно в соединительнотканном слое кожи независимо от хряща. Таким путем развиваются покровные (кожные, или вторичные) кости. Именно так возник в эволюции костных рыб ряд новых элементов скелета, имеющих большое биологическое значение. Не будучи связанными с первичными (исходными) функциями хрящевого скелета, вторичные костные образования открывают возможность большого разнообразия форм, отражающих приспособления к разным условиям жизни. Таковы, например, вторичные челюсти, принимающие на себя хватательную функцию и отличающиеся большим разнообразием строения у разных видов в связи с приспособлением их к различным типам питания, гораздо более многообразным, чем у хрящевых рыб. Только вторичные кости образуют жаберную крышку, имеющую большое значение в активизации дыхания. Вторичный плечевой пояс осуществляет связь первичного пояса с черепом. При характерном для рыб неподвижном соединении черепа с позвоночником такая связь определяет собой более прочную фиксацию пояса и усиление его опорной функции.
В эволюции рыб сначала покровные кости возникали в добавление к хрящевому скелету (состояние, характерное, например, для осетровых рыб), а позднее начался процесс окостенения хряща и образование замещающих костей; одновременно покровные окостенения начали погружаться под кожу, соединяясь с замещающими (хрящевыми) костями.
Во внутреннем строении костистых рыб наиболее бросающаяся в глаза особенность — появление плавательного пузыря — гидростатического органа, увеличивающего «плавучесть» и позволяющего рыбам маневрировать без существенной затраты энергии. У хрящевых рыб это возможно только при движении, что, естественно, требует значительной затраты энергии. Плавательный пузырь выполняет и некоторые добавочные функции: служит резонатором издаваемых рыбой звуков, может служить резервуаром для накопления резервного запаса кислорода (а у некоторых видов — и органом воздушного дыхания) и т. п.
Отсутствие спирального клапана, свойственного хрящевым рыбам, компенсируется у костистых рыб увеличением относительной длины кишечника и развитием у многих видов пилорических придатков, также увеличивающих общую всасывающую поверхность кишечника. Эти преобразования способствуют усилению интенсивности и эффективности пищеварения.
Своеобразно строение мочеполовой системы костистых рыб. Они, как и хрящевые рыбы, обладают мезонефрическими (туловищными) почками с мочеточниками, соответствующими вольфовым каналам. В отличие от хрящевых костистые рыбы имеют мочевой пузырь. Что же касается половых протоков костистых рыб, то они представляют собой особые образования, не гомологичные ни вольфовым, ни мюллеровым каналам. Эти особенности возникают в результате изменений хода эмбрионального развития гонад и, по-видимому, связаны с приспособлением к выведению большого количества половых продуктов; плодовитость костистых рыб значительно выше, чем хрящевых. Впрочем, рассмотренные особенности мочеполовой системы являются специфическим свойством только костистых (и некоторых других костных) рыб и не получили дальнейшего развития в эволюции позвоночных животных.