Реферат: Одноклеточные альтруисты
Однако цитохром с не всегда нужен для запрограммированной смерти. Апоптоз в сердечной ткани, например, вообще протекает без этого фермента, он так и не выходит из межмембранного пространства.
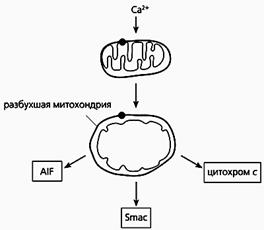
Фрагмент схемы апоптоза, протекающего по митохондриальному пути.
Под действием избытка ионов кальция митохондрия разбухает, через пору из нее выходит цитохром с и два белка - AIF и Smac. Первый белок индуцирует апоптоз, а второй активирует некоторые прокаспазы.
Цитохром с может высвобождаться в ответ на повышение концентрации ионов Са2+, которое вызывает открывание поры. Но выход фермента “на свободу” может и не зависеть от этих ионов, тогда процесс контролируют белки семейства Bcl-2 (B-cell leukaemia-2 - лейкемия В-клеток-2). Именно они регулируют апоптоз на уровне митохондрий. Одни из белков этого большого семейства (Bcl-2, а также Bcl-xL, Bcl-w, Mcl-1, Al и Boo) предотвращают апоптоз; другие (Вах, Bad, Bok, Bcl-xS, Bak, Bid, Bik, Bim, Krk, и Mtd) способствуют самоубийству. Вот один пример работы белков этого семейства регуляторов. Цитозольный белок Bid расщепляется каспазой-8, активируемой через “рецепторы смерти”, и лизосомными протеазами катепсинами, чей выход из лизосом стимулирует эта же каспаза. Образовавшийся активный белок - усеченный Bid (truncated Bid - t-Bid) - изменяет конформацию другого проапоптозного белка, Вах, после чего тот встраивается во внешнюю мембрану митохондрий, где формирует комплекс с порином. Вместе они выстилают канал, по которому из межмембранного пространства выходят цитохром с и проапоптозные белки. Но если в дело вмешивается Bcl-2, действующий как антиоксидант, выход цитохрома блокируется.
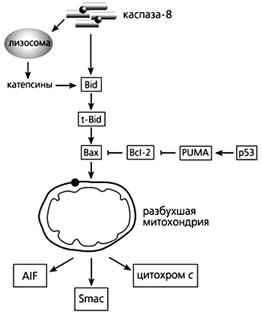
Фрагмент схемы апоптоза, протекающего под контролем белков семейства Bcl-2, а также с участием p53.
Объяснения в тексте.
В запуске апоптоза, вызванного повреждениями ДНК, активацией онкогенов и гипоксией, принимает участие белок-53 (р53), взаимодействуя с Вах, стимулируя “рецепторы смерти” и апоптозные гены. р53 активирует модулятор суицида PUMA (p53 upregulated modulator of APOptosis), который затем связывает Bcl-2 и выводит из строя этот препятствующий апоптозу белок. Тем самым выход цитохрома с из митохондрий уже ничем не сдерживается.
Некоторые белки, связывающие ионы кальция, например ALG-2, кодируемый одноименным геном (APOptosis-linked gene-2), тоже принимают участие в запрограммированной смерти. Так, взаимодействием ALG-2 и белка Alix (ALG-interacting protein X, известный и как AIP1) осуществляется регуляция апоптоза. Часть сложной молекулы ALG-2 представляет собой еще один апоптозный белок кальпоин.
Кальпоины специфически расщепляют белки, содержащие области, называемые PAST-мотивами и богатые остатками четырех аминокислот - пролина (P), аспарагиновой кислоты (A), серина (S) и треонина (T). (Среди этих белков - фермент ДНК-лигаза, у которого такой мотив находится в N-концевой области.) Кроме того, кальпоины освобождают каспазы от связанных с ними ингибиторов апоптозных белков (inhibitors of APOptosis proteins, IAPs).
Впервые обнаруженные у вирусов, гомологи IAP позже были найдены у всех эукариот, от дрожжей до млекопитающих. У последних открыто восемь представителей этого семейства ингибиторов, и каждый имеет от одного до трех 70-аминокислотных участков, называемых BIR-доменами (baculoviruses inhibitor of APOptosis repeat domains - повторяющиеся домены бакуловирусного ингибитора апоптоза). Кодируемый Х-хромосомой IAP (XIAP) и клеточные ингибиторы апоптозных белков (cellular inhibitors of APOptosis protein; cIAP1, 2) содержат по три N-концевых BIR-домена и С-концевой RING-домен, кодируемый геном ring (really interesting new gene - действительно интересный новый ген). Эти белки ингибируют каспазы-3, -7, -9: BIR-домены целенаправленно связываются с ними, блокируя их активные центры; RING-домен обеспечивает деградацию каспаз, пришивая к ним “черную метку” - 76-аминокислотный белок убиквитин. Присоединившие его белки подвергаются расщеплению протеасомой *.
* Об этом специфическом пути деградации белков см.: Абрамова Е.Б., Карпов В.Л. Протеасома: разрушение во имя созидания // Природа. 2003. №7. С.36-45.
Есть и другой эндогенный ингибитор каспаз - FLIP (FLICE-inhibitory protein). Он взаимодействует с пособниками суицида - сигнальными комплексами, блокируя активацию прокаспазы-8 и передачу проапоптического сигнала от “рецепторов смерти”.
Как все ингибиторы протеаз, XIAP, cIAP1 и cIAP2 сами подвержены отрицательной регуляции. В частности, митохондриальный полипептид Smac/DIABLO (direct IAP-binding protein with low pI - белок с низкой pI, прямо связывающий IAP), выходя из митохондрий в цитозоль, связывается с ингибиторами XIAP, cIAP1 и cIAP2 и мешает им взаимодействовать с каспазами. Но XIAP, cIAP1 и 2 способны противостоять ингибирующему действию Smac/DIABLO, присоединяя к нему убиквитин - сигнал для деградации. Одна из изоформ Smac/DIABLO, Smac3, сама стимулирует присоединение убиквитина к XIAP. В высоких концентрациях каспазы-3 и -7 могут расщеплять XIAP и без посредников.
Как видим, клетка никогда не бросается в апоптоз без “раздумий”. Она сначала задает себе гамлетовский вопрос: “Быть или не быть?”. Ответ на него зависит от соотношения про- и антиапоптозных белков; между ними идет борьба, и побеждает та сторона, которая успевает навесить на другую “черную метку” - убиквитин. А протеазам совершенно все равно, на ком эта метка - они беспристрастно расщепляют меченые белки. Таким образом, выбор клетки между жизнью и смертью зависит не столько от внешних обстоятельств, сколько от ее собственного внутреннего состояния. Даже если организм приказывает клетке погибнуть, она подчиняется ему только в том случае, когда сама расположена к такому исходу. Бывают мутанты, которые категорически не хотят умирать, при том что все внешние обстоятельства побуждают их к суициду - как правило, это раковые клетки.
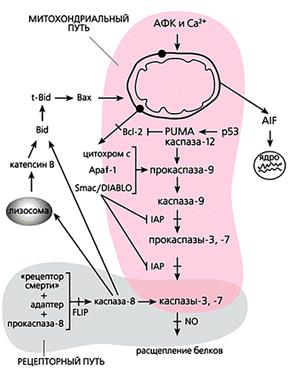
Общая схема “классического” апоптоза млекопитающих.
Мы перечислили все виды орудий самоубийства, которыми клетка многоклеточного организма пользуется, когда он приказывает и когда она сама того хочет. Но следует понимать, что их действие неоднозначно. Например, кальций и активные формы кислорода участвуют во всех без исключения процессах жизнедеятельности как вторичные посредники в передаче сигналов и регулируют все функции клетки: размножение, дыхание, обмен веществ, движение, рост. Тот же кальций способен, в частности, активировать антиоксидантные ферменты, защищающие клетку от окислительного стресса. АФК в микромолярных концентрациях необходимы для пролиферации клеток, их подвижности и хеморецепции; более того, некоторые виды (например, NO) даже блокируют апоптоз посредством избирательного нитрозилирования эффекторных каспаз. Да и каспазы тоже могут выполнять вполне “мирные” функции - управлять дифференцировкой некоторых типов клеток.
Таким образом, лишь немногие из орудий самоубийства служат только этой цели: большинством из них клетка пользуется и в “домашнем быту”. Возникает вопрос: а все ли они необходимы для совершения суицида? И если нет, то зачем производить столь непомерный арсенал орудий? На эти вопросы мы сможем ответить, когда рассмотрим специфику протекания запрограммированной смерти у одноклеточных организмов.
Суицид у одноклеточных
Практически у всех простейших организмов внешние признаки апоптоза те же самые, что у клеток многоклеточных, и, как у них, к гибели приводят разные причины. У одноклеточных водорослей ее вызывает отсутствие света или окислительный стресс, а предотвращают ингибиторы каспаз и антиоксидантный фермент, расщепляющий Н2О2, - каталаза (каспазы у них тоже выявлены - с помощью антител). Паразитические жгутиконосцы кончают жизнь самоубийством, если им помогают ингибитор протеинкиназы С ставроспорин (он же стимулирует апоптоз и в клетках млекопитающих) и перекись водорода, а предотвращают тоже ингибиторы каспаз. Экстракт цитоплазмы паразитического простейшего Leishmania major в опытах вызывал характерные для апоптоза изменения в изолированных ядрах млекопитающих, свидетельствуя о присутствии в нем AIF-подобных белков. Изолированные митохондрии L.major теряли цитохром c при инкубации с рекомбинантным Вах человека даже тогда, когда Вах был лишен трансмембранного домена, необходимого для встраивания в наружные мембраны митохондрий. Следовательно, у L.major есть белки, способные взаимодействовать с Вах.
У дрожжей найдены практически все орудия суицида, имеющиеся у многоклеточных, кроме лизосомальных катепсинов и представителей семейства Bcl-2. Тем не менее, гены проапоптозных белков млекопитающих при экспрессии в дрожжах вызывают их запрограммированную гибель, а Bcl-2 защищает пекарские дрожжи (Saccharomyces сerevisiae) от гибели, вызванной окислительным стрессом. У Shizosaccharomyces pombe найден Rad9-белок, содержащий последовательность аминокислот, необходимую для взаимодействия с Bcl-2.
К апоптозу дрожжей приводят вещества, которые они сами же и продуцируют: например, уксусная кислота (продукт брожения), перекись водорода или a-фактор - пептидный половой феромон, вырабатываемый a-типом гаплоидных клеток S.сerevisiae. При концентрации <1 мкМ этот феромон стимулирует конъюгацию клеток противоположного типа спаривания с дрожжами a-типа, а при более высокой - вызывает блокаду клеточного цикла и апоптоз. Мутация активируемой феромоном протеинкиназы предотвращает и смерть, и появление ее маркеров.
Слизистые грибы (миксомицеты) - уникальные организмы, проходящие в своем развитии одно- и многоклеточную стадии. Когда в почве достаточно питательных веществ, миксомицеты существуют в виде отдельных амебообразных клеток - миксамеб. Во время голодания они сползаются и образуют одно большое клеточное скопление - плазмодий. Некоторое время он ползает по субстрату, а потом останавливается и начинает формировать плодовое тело. Часть его клеток при этом превращается в споры, а часть - в стебельковые клетки, выполняющие опорную функцию. Именно они подвергаются запрограммированной гибели, в целом похожей на классический апоптоз, но сопровождающейся интенсивным появлением вакуолей. Интересно, что ДНК стебельковых клеток не распадается на фрагменты. Ингибиторы каспаз не препятствуют гибели клеток, но нарушают нормальное развитие плодовых тел. У амеб Dictyostelium discoideum найдены гомологи четырех белков, участвующих в запрограммированной смерти клеток многоклеточных организмов: AIF, Alix и два ALG-2. Примечательно, что гены обоих гомологов ALG-2 экспрессируются в растущих миксамебах. Мутанты, лишенные одного или обоих этих генов, формируют нормальные плодовые тела, однако мутанты по гену alix имеют ярко выраженные дефекты развития.
У инфузорий есть все виды орудий самоубийства (в том числе каспазы-8 и -9, но не -3), кроме белков семейства Bcl-2 (пока их просто не искали). Инфузория Paramecium tetraurelia синтезирует катепсины L и S. У Tetrahymena pyriformis, штамма W, найден катепсин В.
Таким образом, как мы видим на примере самых примитивных простейших, для запуска апоптоза в принципе достаточно двух компонентов: активной формы кислорода и хотя бы одной цистеиновой протеазы, т.е. каспазы. В этой связи отметим, что инициаторные каспазы в ходе эволюции возникают раньше, чем эффекторные; последние не найдены ни у кого из простейших. У большинства эукариот ведущая роль в стимуляции апоптоза принадлежит митохондриальным факторам - цитохрому с и AIF. Тем не менее, не имеющий митохондрий паразит мочеполовой системы человека Trichomonas vaginalis каким-то образом обходится без обоих факторов. Отсутствие гомологов ALG-2 у слизистых грибов не нарушает образования плодовых тел, из чего следует, что и эти белки необязательны для апоптозного процесса. Следовательно, большинство белков, принимающих участие в запрограммированной смерти, предназначено для более тщательной ее регуляции. А тщательность вряд ли была бы возможна при малом количестве звеньев.
* * *
Биологический смысл существования сразу нескольких систем запуска апоптоза, по-видимому, в том, чтобы его могли независимо друг от друга побуждать стимулы разного рода: повреждение генома, стрессовые воздействия, необходимость для организма в целом и т.д. Примитивным одноклеточным организмам просто не нужна столь многокомпонентная система стимуляции апоптоза, они гибнут только при стрессовых воздействиях, потенциально опасных для генома и, следовательно, для популяции в целом. Многоклеточному же организму необходимо, чтобы клетки
- были всегда готовы покончить с собой по его приказу там, где это потребуется;
- сами принимали такое решение при возникновении неполадок в собственном геноме или при угрозе их появления.
Для этого и нужно, чтобы апоптоз могли запустить и специальные сигнальные молекулы (через “рецепторы смерти”), и повышение уровней АФК и цитоплазматического кальция (всегда сопровождающее любые стрессы), и повреждение генома.