Реферат: Поведение чайки
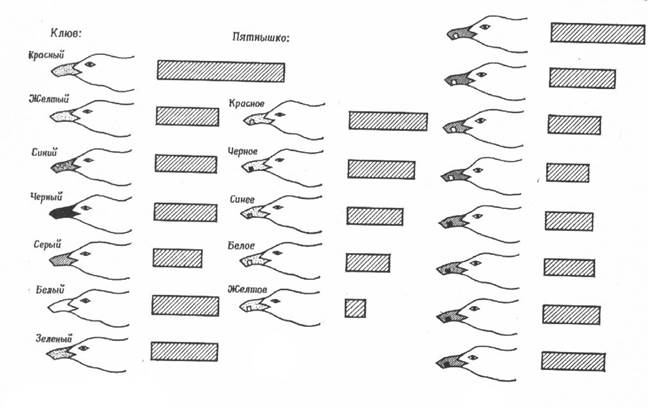 расположенный клюв взрослой птицы часто движется горизонтально перед глазами птенца.
расположенный клюв взрослой птицы часто движется горизонтально перед глазами птенца.
Дальнейший анализ результатов показал, что вертикально движущийся вертикальный стимул вызывал не больше реакций, чем неподвижный вертикальный. Кроме того, и горизонтальное, и вертикальное перемещение горизонтального стимула было одинаково действенно и, как это ни странно, превосходило по эффективности неподвижный горизонтальный стимул. Наиболее осторожная интерпретация этих результатов может состоять в том, что адекватными стимулами для пищевой реакции птенцов служат два типа движения: 1) горизонтальное перемещение и 2) движение в направлении, перпендикулярном продольной оси стимула. Это позволяет объяснить, почему неподвижная вертикальная палочка оказывается не менее эффективным стимулом, чем вертикальная палочка, движущаяся вертикально. Дело в том, что в обоих случаях отсутствуют движения указанных типов. Однако в случае горизонтальной палочки вертикальное движение перпендикулярно продольной оси стимула и потому так же эффективно, как горизонтальное, и реакция здесь при обоих движениях более интенсивна, чем при полной неподвижности стимула.
На следующем этапе мы испытывали вертикальные стимулы различных диаметров, и пять скоростей движения. Палочка диаметром около 8 мм была наиболее эффективна вне зависимости от скорости движения. Скоростью, вызывающей наиболее интенсивную реакцию независимо от диаметра палочки, оказалась скорость 12 см/с. Эти результаты показывают, насколько точно реактивность в процессе эволюции подгоняется к параметрам стимулов. Толщина клюва взрослой птицы в вертикальном направлении 10,6 мм, а в горизонтальном-3,1 мм, так что средняя толщина составляет как раз около 8 мм. Кроме того, как показал анализ фильмов, снятых в естественных условиях, средняя скорость движения клюва при кормлении равна около 14,5 см/с.
Проведенный недавно эксперимент добавил еще одну особенность к образу идеального для птенцов стимула. Вертикальная палочка, находящаяся выше уровня глаз птенца, гораздо эффективнее, чем такой же стимул, предъявленный внизу. Кроме того, предпочтение отдается наклонным объектам. Такая избирательность уменьшает вероятность возникновения у птенца реакции на ноги родителей, расположенные, естественно, ниже уровня его глаз.
Теперь понятно, по крайней мере, в общих чертах, каким образом только что вылупившийся птенец отличает клюв родителей от других внешних объектов. На следующем этапе нам предстояло выяснить, изменяется ли восприятие птенца в первые несколько дней жизни. Предъявляли те же три набора по пять моделей в каждом семидневным птенцам и нашли, что их поведение заметно отличается от поведения только что вылупившихся птенцов. Более взрослые птенцы оказались чувствительными даже к незначительным изменениям формы головы и клюва. Кроме того, они четко отличали модели собственных родителей – озёрных чаек - от моделей серебристых чаек.
Итак, данные указывают на то, что недавно вылупившийся птенец лучше всего реагирует на очень простые стимулы. Хотя экспериментатор может создать модель, в качестве стимула значительно более эффективную, чем настоящая птица, в естественных условиях признаки родителей точнее соответствуют «образу» стимула для пищевой реакции, чем особенности любого другого внешнего объекта. Кроме того, по мере выкармливания птенца родителями их образ в памяти птенца детализируется. Семидневные птенцы клюют только модели достаточно сходные с взрослыми птицами того же вида..
Полагается, что вопрос о том, остаются ли все элементы стимула действительно идентичными. требует дополнительного изучения. В экспериментах Тинбергена и Пердека модель держали в руке, поэтому при ее маятникообразном движении вместе с кистью руки пятно на лбу двигалось медленнее и по более короткой дуге, чем на клюве. Кроме того, птенцу приходилось тянуться вверх, чтобы клюнуть пятно на лбу.
Можно повторить опыты Тинбергена - Пердека, но добавить еще одну модель, пятно у которой располагалось на лбу, но она закреплялась на стержне таким образом, что пятно оказывалось на таком же расстоянии от оси вращения, как и пятно на клюве у другой модели. Кроме того, экспериментальная установка была снабжена подвижным полом, высоту которого подбирали так, чтобы красное пятно на модели всегда оказывалось на уровне глаз птенца. Эту третью модель назвали «быстрой», так как пятно у нее на лбу перемещалось быстрее, чем у «медленной» модели Тинбергена и Пердека. Если гипотеза о роли скорости движения верна, то «быстрая» модель с пятном на лбу должна быть так же эффективна, как и модель с пятном на клюве.
Результаты оказались совершенно однозначными. Только что вылупившиеся птенцы одинаково охотно клевали и «быструю» модель с пятном на лбу, и обычную модель с пятном на клюве. Затем птенцы были возвращены в гнездо и подвергнуты повторным тестам на 3-й и 7-й дни жизни. Как и предполагалось, реакция на модель с пятном на клюве постепенно улучшалась по сравнению с обоими вариантами модели с пятном на лбу.
Классическая трактовка экспериментов Тинбергена - Пердека сводилась к тому, что существует некий врожденный механизм, включающийся, как полагали, при восприятии определенных довольно сложных стимулов. Эксперименты показали, что пищевое поведение птенцов чайки вначале определяется сравнительно простыми стимулами, но позже в результате научения используются более сложные стимулы. Эксперименты указывают, видимо, на то, что необходимо заново исследовать и другие случаи, которые считались примерами действия врожденных «запускающих» механизмов.
Результаты исследований подсказывают также новую интерпретацию еще одного представления, широко используемого исследователями поведения - представления о классическом условном рефлексе. В общеизвестных экспериментах И. П. Павлова до начала или во время действия безусловного стимула, вызывающего биологически адекватную реакцию, животному предъявлялся новый для него условный стимул. После некоторого числа таких сочетаний животное начинало отвечать и на отдельно предъявляемый условный стимул. В классических экспериментах Павлова звук звонка подкреплялся пищей. Через некоторое время в ответ на воздействие одного лишь звонка у животного можно было наблюдать выделение слюны.
Психологов давно интересовало, насколько полезной в обычной жизни животного может быть такая способность устанавливать связь между стимулами различных модальностей. Как в эволюции могла выработаться способность к такой форме научения, которая в естественных условиях, по-видимому, так мало используется? Результаты наводят на мысль, которую стоило бы проверить. Вспомним о том, что вначале птенец реагирует на самые простые признаки клюва взрослой птицы (безусловные стимулы), но при этом одновременно воспринимает и множество других деталей головы (условные стимулы). Под влиянием пищевого подкрепления эффективным стимулом для клевания становится все более детализированный зрительный образ.
Такого рода процесс, который можно назвать «обострением восприятия», отличается от классического условного рефлекса, наблюдаемого в лаборатории, тем, что условный и безусловный стимулы здесь физически идентичны. Не исключено, что способность к выработке классических условных рефлексов сформировалась в эволюции главным образом как механизм, лежащий в основе феномена обострения восприятия. В этом случае исследователи классических условных рефлексов, по существу, изучают побочные эффекты этого механизма. Но сейчас эти соображения - не более чем одна из возможных гипотез.
Изучали и многие другие аспекты пищевого поведения, и об одном вопросе необходимо упомянуть: речь идет о распознавании пиши. Узнают ли только что вылупившиеся птенцы пищу, когда сталкиваются с нею? Чтобы выяснить это, поставили четыре чашечки с кормом в углах небольшого ящика и пронаблюдали, как выведенные в инкубаторе птенцы первый раз в жизни находят пищу. Время, затраченное птенцом на поиски пищи, оказалось обратно пропорциональным частоте клевков. Этого и следовало ожидать, если исходить из того, что птенцы разыскивают пищу только методом проб и ошибок.
Если птенцам дать насытиться и пересадить их в другое место, пока они не проголодаются, то в повторном опыте они находят пищу гораздо быстрее. В третьем опыте необходимое время сокращается до минимума. Это нельзя объяснить увеличением частоты клевков, так как во втором в последующих опытах она лишь незначительно выше, чем в первом. Следовательно, птенцы очень быстро научаются распознавать пищу или по крайней мере ее местонахождение.
Если вылупившийся птенец вначале не умеет распознавать пищу, значит ли это, что он вынужден полагаться только на метод проб и ошибок, пока, случайно наткнувшись на корм, он не начнет быстро обучаться поискам? Наблюдения и эксперименты показывают, что есть несколько механизмов, помогающих птенцу ускорить первый поиск пиши. Запомним, что если птенец не клюет пищу, то взрослая птица обычно берет ее в клюв. При первом кормлении птенец продолжает клевать клюв родителя и после того, как срыгнутая пища падает на дно гнезда. В конце юнцов взрослая птица подбирает пищу, и птенец натыкается на нее при клевке, нацеленном на клюв родителя. Это наводит на мысль об адаптивном научении малой точности клевков только что вылупившегося птенца, так как промахи повышают вероятность того, что он при этом случайно натолкнется на пищу.
Другой механизм, помогающий птенцу быстрее научиться распознавать пищу, связан с присутствием в гнезде других птенцов. Клевки одного птенца часто бывают нацелены на белый кончик клюва другого. Обычно птенцы вылупляются с интервалом около 12 часов, и к моменту вылупления следующего птенца старший уже получает корм. Если младший пытается во время такой кормежки клевать клюв старшего, то он тоже может наткнуться на пищу. Наблюдения за гнездами подтвердили, что иногда птенцы впервые находят корм именно таким способом.
Процесс распознавания корма изучали, используя три группы птенцов серебристой чайки, выведенных в инкубаторе. Птенцы контрольной группы помещались в коробки с пищей поодиночке. Птенцы двух экспериментальных групп размещались попарно, но в одном случае второй птенец тоже никогда не видел пищи, а во втором в качестве пары подбирался птенец, ранее кормившийся в коробке. Оказалось, что птенцы, тестируемые поодиночке, дольше всех не находили пищи. Несколько меньше времени на поиск корма требовалось птенцу, компаньон которого был так же неопытен, как и он сам. Быстрее всех с задачей справлялся птенец, находившийся в коробке вместе с птенцом, уже умеющим распознавать пищу.
Такие различия нельзя объяснить только более активным «исследовательским» клеванием по принципу проб и ошибок в присутствии компаньона, так как клюв последнего, наоборот, отвлекал тестируемого птенца от таких проб. Присутствие неопытного птенца побуждало другого больше передвигаться в ящике, и это приводило к более быстрому, чем у одиночных птенцов, обнаружению пищи. Влияние обученного птенца этим не ограничивалось, так как все неопытные птенцы в его присутствии впервые находили пищу, целясь в клюв кормящегося компаньона.
Птенец чайки вступает в жизнь, обладая недостаточно координированной, неточной реакцией клевания, определяемой такими простыми признаками стимула, как форма и характер движения (мотивацией служит голод, источниками стимула - родители и другие птенцы в гнезде). Птенец вначале не способен распознавать пищу, но, промахиваясь при клевках, нацеленных на клюв другой особи, он натыкается на корм и быстро научается узнавать его. Благодаря пищевому подкреплению птенец запоминает облик родителей. В результате практики повышается точность клевков и совершенствуется оценка расстояния до цели. Птенец научается также поворачивать голову при выпрашивании пищи, что ведет к появлению отличия этой реакции от клевков, направленных непосредственно на пищу.
Обнаруженная картина позволяет думать, что и развитие других инстинктов включает элемент научения. Для формирования стереотипной видоспецифической формы поведения важно только, чтобы процесс научения у всех особей данного вида был очень сходным. На описанном примере с чайками ясно видно, что поведение нельзя осмысленно расчленить на врожденные и приобретенные формы, так же как невозможно выделить какую-либо часть поведенческого репертуара, связанную только с научением. Развитие поведения - это мозаика, обусловленная непрерывным взаимодействием организма и среды.
Источники: литература
«Жизнь животных» том 5, М. «Просвещение» - 1969
«Птицы» М. «Мир», 1983
К. Вили, В.Детье «Биология» М. «Мир», 1974
Р. Хайнд «Поведение животных» М. «Мир», 1975
«Биологический Энциклопедический Словарь», М. «Советская энциклопедия» - 1989
«Большой Энциклопедический Словарь», М. «Большая Российская Энциклопедия» - 2002