Реферат: Рост аксона
Кончик растущего аксона удлиняется, образуя конус роста. Рамон-и-Кахаль первым обнаружил, что конус роста является частью аксона, ответственной за навигацию и удлинение его в направлении конечной цели . Конус роста удлиняется и сокращается за счет широких пластинок, называемых ламмелиподиями, и тонких, остроконечных выпячиваний, называемых филоподиями Филоподий достигают размеров в несколько десятков микрометров и могут удлиняться и сокращаться, как бы ощупывая субстрат во всех направлениях. Филоподий адгезируют к определенному субстрату и тянут конус роста в этом направлении.
Актин играет ключевую роль в подвижности конуса роста (рис. 1) . Как ламеллиподии, так и филоподий богаты филаментным актином и веществами, ингибирующими полимеризацию актина, такими как грибной токсин цитохалазин В, обездвиживающий конусы роста. Выпячивание и ретракция ламеллиподий и филоподий, а также движение вперед самого конуса роста, скорее всего, управляется двумя процессами: (1) полимеризацией и разборкой актиновых филаментов и (2) зависимой от миозина транслокацией актиновых филаментов прочь от ведущего края конуса роста. Оба процесса используют энергию гидролиза АТФ и могут управляться белками, связывающими актин. Кальций, протеинкиназы и другие внутриклеточные вторичные посредники управляют активностью актин-связывающих белков. Например, остановка роста и ретракция конуса роста, два частых события в процессе роста аксонов, связаны с входящим током кальция и увеличением частоты кратковременных повышений концентрации кальция в цитоплазме клетки.
|
|
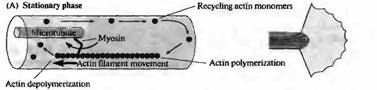
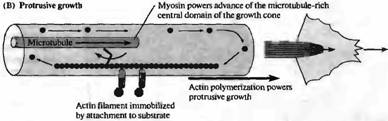
| Рис. 1. Модель подвижности конусов роста на основе актина. (А) Поперечный срез (слева) и вид сверху (справа) на конус роста в стационарной фазе. Миозин, связанный с микротрубочками, обеспечивает движение актиновых филаментов в направлении назад, в то время как в филаментах постоянно происходит процесс полимеризации со стороны ведущего края конуса роста и деполимеризация в центре. (В) Аналогичные изображения конуса роста во время роста. Актиновые филаменты иммобилизировались присоединением к субстрату. Полимеризация актина в этом случае приводит к продвижению конуса роста, в то время как миозин обеспечивает движение микротрубочек вперед, продвигая центральный домен конуса роста. |
Молекулы адгезии клетки и внеклеточного матрикса и рост аксона
Молекулы клеточной адгезии управляют ростом аксона, обеспечивая наиболее благоприятное окружение для вытягивания конуса роста. Клеточные молекулы адгезии представляют собой трансмембранные или связанные с мембраной гликопротеины, характеризующиеся структурными мотивами своих внеклеточных частей, которые во многом гомологичны постоянным доменам иммуноглобулинов и фибронектина 3 типа (рис.2). Представителями надсемейства этих иммуноглобулинов (Ig) являются клеточные молекулы адгезии (N-CAM), молекулы адгезии нейроглии САМ (NgCAM), TAG-1, MAG, и DCC83) . Эти молекулы обеспечивают адгезию клеток друг к другу посредством гетерофильных связей между различными надсемействами иммуноглобулинов (например, связь между NrCAM и TAG-1). Дополнительно присутствующей везде молекулой клеточной адгезии является молекула N-кадгерина (N-cadherin, рис. 2), которая обеспечивает кальций-зависимую адгезию между клетками.
В культуре клеток экспрессия N-CAM и N-кадгерина в клетках приводит к их агрегации, вытягиванию аксонов в сторону клеточных субстратов, но не субстратов внеклеточного матрикса, а также соединению отдельных растущих аксонов в пучки (фасцикулы, fascicles). Стимуляция роста аксонов молекулами клеточной адгезии не обеспечивается просто «липкостью» субстрата; она управляется активацией рецепторов, связанных с тиро1инкиназой, например рецептора к фактору роста фибробластов (fibroblastgrowthfactor, FGF).
| Рис. 2. Два класса молекул адгезии нервных клеток. (А) N-кадгерин способствует гомофильной кальций-зависимой адгезии. (В) Члены надсемейства иммуноглобулинов характеризуются многочисленными повторами связанных друг с другом дисульфидными мостиками циклов, которые гомологичны доменам, впервые описанным в постоянной части молекул иммуноглобулина. Многие из этих молекул клеточной адгезии также содержат несколько доменов, сходных с повторами в фибронектине III типа (показано в виде прямоугольников). У различных видов животных имеется большое количество гомологичных белков со сходными именами. | 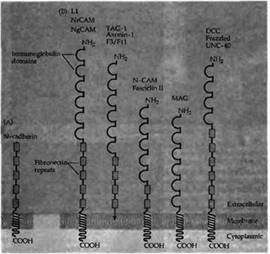 |
Рецептор FGF включает внутриклеточный каскад, связанный с фосфорилированием тирозина, что приводит к удлинению аксона. Белковая тирозинфосфатаза, фермент, удаляющий остатки фосфата с тирозина, также участвует в регулировании этих сигналов. Молекулы адгезии внеклеточного матрикса, включая ламинин, фибронектин, тенасцин (Л, цитотактин), а также перлецан, тоже являются благоприятным субстратом для роста нервных отростков85 · 86) . Эти гликопротеины с большой массой имеют две или более сходные субъединицы, удерживаемые вместе благодаря дисульфидным мостикам (рис. 2). Каждая субъединица характеризуется повторяющимися структурными мотивами. Белки внеклеточного матрикса взаимодействуют с клетками через семейство рецепторов, называемых интегринами. Было идентифицировано большое количество изоформ субъединиц интегрина. Каждая комбинация приводит к формированию рецепторов с различными свойствами. Интегрины обеспечивают структурные связи между белками внеклеточного матрикса и внутриклеточным актиновым цитоскелетом, регулируя форму клетки и ее миграцию. Кроме того, они активируют внутриклеточные сигнальные пути, которые управляют ростом клетки, пролиферацией и дифференцировкой.
Исследования при помощи специальных блокирующих антител показывают, что конусы роста редко используют только один субстрат для своего движения; несколько типов молекул адгезии клетки и внеклеточного матрикса могут обеспечивать рост нервных отростков у определенных типов нейронов. Например, для полного ингибирования роста аксонов в сторону шванновской клетки необходимо применять одновременно антитела к Ll/NgCAM, N-кадгерину и интегринам. Один тип антител сам по себе не может помешать росту аксона.
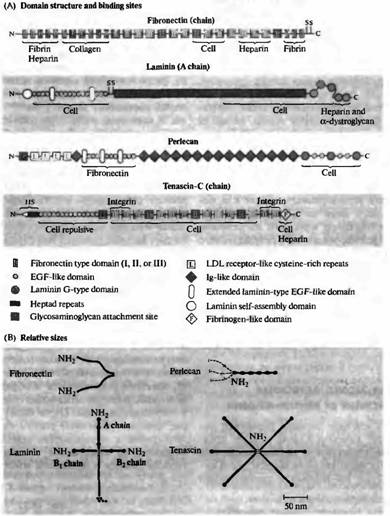 |
| Рис. 3. Белки внеклеточного матрикса, опосредующие адгезию клеток к субстрату и отталкивание. (А) Схематическое представление доменной структуры цепей, состоящей из фибронектина, ламинина (А цепь), перцелана и тенасцина-С, а также участков, к которым присоединяются клетки, коллаген, гепарин, фибронектин, интегрин, фибрин и альфа-дистрогликан. S указывает на расположения остатков цистеина, которые образуют дисульфидные мостики между цепочками. Тенасцин-С содержит как связывающие, так и отталкивающие домены; основной его эффект, однако, отталкивание клеток и конусов роста. (В) Молекулы, изображенные в определенном масштабе. Все, кроме перцелана, являются олигопротеинами, цепи связаны друг с другом дисульфидными мостиками. Пунктирной линией показан гепарансульфат 90 и 170 нм длиной, связанный с N терминальным доменом перцелана. (ИзEngel Л., 1992.) |
Управление ростом аксона
Аксоны нервных клеток могут достигать 1 метра и более в длину, образуя синапсы в определенном месте на строго определенной клетке в области, где имеется большое количество других потенциальных клеток-мишеней. Две основные теории, касающиеся того, каким образом устанавливается такая специфичность в синаптических связях во время развития, были предложены в первой четверти двадцатого века. Согласно одной, нейроны и их мишени заранее запрограммированы на образование определенных синаптических связей. Другая теория считает, что первоначально связи устанавливаются более или менее случайно, а затем часть из них реорганизовывается благодаря влияниям на нейроны со стороны клеток-мишеней, в результате чего происходит устранение ошибочных синапсов и гибель неправильно соединенных клеток. Экспериментальные доказательства, полученные к настоящему времени, говорят в пользу того, что рост аксонов и образование синаптических связей является селективным процессом; аксон направляется строго к своей цели благодаря определенным сигналам в своем окружении.
Навигация аксона, зависящая и не зависящая от клетки-мишени
Какие внеклеточные сигналы управляют конусом роста? Рамон-и-Кахаль сначала предложил хемоаттрактантную модель управления аксоном, согласно которой конусы роста аксонов следуют по градиенту концентрации некоторых молекул, вырабатываемых клетками-мишенями. Такой механизм возможен для роста аксона, когда расстояние между телом нейрона и его мишенью очень короткое. Например, Лумсден и Дэвис изучали рост аксонов из ганглия тройничного нерва, расположенного в голове мыши, в соседние эпителиальные ткани, расстояние до которых составляло менее 1 мм (эти аксоны в конечном счете образовывали сенсорную иннервацию вибрисс, Если развивающийся ганглий тройничного нерва помещался в культуру вблизи эксплантантов из нескольких периферических тканей, нервные отростки росли из ганглия в направлении собственной клетки-мишени, игнорируя другие ткани. Эксплантаты эпителия ткани-мишени оказывали подобное действие на рост аксонов только в том случае, если они брались у эмбриона в период становления иннервации.
В противоположность этому, способность аксонов спинальных мотонейронов расти в направлении конечностей не зависит от наличия мышечной клетки-мишени. Это было показано путем удаления в раннем периоде сомита, из которого развивается мускулатура конечностей. Аксоны мотонейронов направляются нормально из спинного мозга, врастают в конечность и образуют определенный паттерн мышечных нервов, даже в отсутствие мышцы. Таким образом, факторы, которые управляют ростом аксонов мотонейронов к определенной мишени на конечности, не выделяются мышцами, с которыми аксоны в конечном итоге образуют связи.
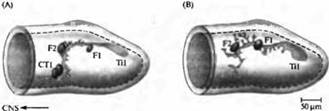
| Рис. 4. Конусы роста периферических нейронов используют клетки-ориентиры для навигации в конечностях кузнечика. (А) В нормальном эмбрионе аксоны Ti1 нейрона встречают на своем пути в центральную нервную систему серию клеток-ориентиров: клетки Fl, F2 и две клетки СТ1. (В) Если СТ1 клетку убить на ранней стадии развития, нейрон Ti1 образует несколько аксональных веточек из точки, где находится F2, и конусы роста направляются в неправильном направлении. |
|
Навигация по клеткам-ориентирам (guidepost cells)
Когда дистанция от нейрона до его цели составляет больше чем несколько сотен микрон, его путь обозначен специальными промежуточными целями. Например, конус роста, идущий от сенсорной клетки конечностей у развивающегося кузнечика, делает несколько резких поворотов на своем пути в ЦНС (рис. 4). Эти повороты происходят в тот момент, когда конус роста касается так называемых клеток-ориентиров (guidepost cells). Такое поведение указывает на наличие взаимодействия с клетками-ориентирами, ответственными за перенаправление конусов роста. Этими клетками чаше всего являются незрелые нейроны. Эти взаимодействия можно продемонстрировать при помощи удаления клеток-ориентиров лучом лазера до того, как их достигнет конус роста. В этом случае не происходит соответствующего изменения в траектории движения конуса роста.
Синаптические взаимодействия с клетками-ориентирами
В некоторых случаях аксоны образуют кратковременные синаптические контакты с клетками-ориентирами во время развития. В развивающемся гиппокампе, например, аксоны из энторинальной коры сначала образуют синапсы с временной популяцией нейронов, клетками Кахаля—Ретциуса. Позднее, по мере появления гранулярных клеток и их созревания, энторинальные аксоны покидают клетки Кахаля—Ретциуса и образуют связи с дендритами гранулярных клеток. После этого клетки Кахаля—Ретциуса исчезают. В другом примере аксоны нейронов из ядра ЛКТ в зрительной системе млекопитающих достигают развивающуюся корковую пластинку раньше, чем образуются их синаптические мишени — пирамидные клетки слоя 4. Поэтому аксоны ядра ЛКТ образуют синапсы с нейронами подпластинки , которые образуются в раннем эмбриогенезе. Нейроны подпластинки лежат под развивающейся корковой пластинкой, и им суждено исчезнуть вскоре после рождения. Через нескольких недель, когда пирамидные клетки слоя 4 достигают своего месторасположения в коре, аксоны из ЛКТ разрывают свои связи с нейронами подпластинки и направляются в кору, чтобы образовать связи, характерные для взрослого животного. Если нейроны подпластинки удалить в раннем периоде развития местной аппликацией нейротоксинов, аксоны ядра ЛКТ прорастают за пределы развивающейся зрительной коры и не могут образовать синаптических связей со своими мишенями.
Механизмы управления аксоном
Молекулы, которые управляют конусом роста, действуют четырьмя основными путями: как аттрактанты или репелленты, на короткой или длинной дистанции.
Некоторые коротко-дистантные сигналы управления аксоном обеспечиваются посредством контакта конуса роста с клеточной поверхностью или с белками адгезии внеклеточного матрикса, которые были описаны ранее в этой главе как промоторы роста аксона. Молекулы адгезии и их рецепторы могут также играть ведущую роль в связывании отдельных аксонов в пучки (так называемая фасцикуляция). Характер роста аксона и иннервация клетки-мишени in vivo и в культуре клеток нарушалась при изменении активности специфических молекул адгезии или их рецепторов, в генетических экспериментах с выключением определенных генов, а также при добавлении определенных типов антител.
Другое влияние молекул внеклеточного матрикса на растущий аксон показано в экспериментах на отдельных клетках, выделенных из ЦНС пиявки и растущих в культуре. Субстраты, которые содержат тенасцин или ламиннн, способствуют не только быстрому образованию нервных отростков у нейронов пиявки, но также влияют на характер роста этих отростков и на распределение кальциевых каналов в клетке. Различные нейроны по-разному отвечают на определенные молекулы внеклеточного матрикса. Таким образом, возникает экономная схема, когда несколько молекул адгезии могут обеспечивать разнообразные эффекты.
|
Рис. 5. Молекулы внеклеточного матрикса определяют паттерн роста нервных отростков в культуре нейронов пиявки. Одиночный нейрон выращен в клеточной культуре на субстрате определенного паттерна. С левой стороны край чашки был покрыт конканавалином A (Con A), а с правой стороны экстрактом, содержащим внеклеточный матрикс (ЕСМ) из глиальных капсул ганглиев пиявки. | 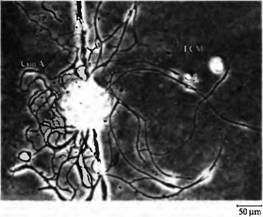 |
Все молекулы адгезии либо способствуют росту, либо нет. Управление ростом аксона на большом расстоянии включает в себя движение аксона вдоль градиента концентрации растворимого фактора.
Навигация конусов роста в спинном мозге
Хороший пример разнообразия механизмов и молекул, которые используют конусы роста для навигации к своим клеткам-мишеням, представляют аксоны комиссуральных интернейронов спинного мозга позвоночных В раннем периоде развития комиссуралъные интернейроны, которые лежат в дорзальной части спинного мозга, «выпускают» аксоны, которые начинают расти в вентральном направлении, пересекая среднюю линию, и затем растут вдоль спинного мозга по направлению к своим синаптическим мишеням (рис. 6) .
Аксоны комиссуральных нейронов первоначально привлекаются к вентральной средней линии белком нетрин-1, растворимым хемоаттрактантом, образуемым специальными клетками базальной пластинки, лежащими вдоль средней линии спинного мозга (рис. 23.24А)111) . Нетрин-1 взаимодействует с рецептором, экспрессируемым комиссуральными нейронами, называемым DCC (который уже упоминался ранее из-за своего взаимодействия с молекулами клеточной адгезии. Существование растворимого фактора, образующегося в базальной пластинке, который способен привлекать (attract) аксоны комиссуральных нейронов, было впервые показано при культивировании кусочков дорзальной части спинного мозга изолированно, либо вместе с кусочками базальной пластинки (рис. 7) . Аксоны комиссуральных нейронов растут четко по направлению к базальной пластинке, даже если эксплантаты находятся друг от друга на расстоянии нескольких сотен микрон. Эта дистанция слишком велика для того, чтобы ее прошел филоподий конуса роста и требует растворимого фактора. Такой фактор, нетрин-1, входящий в семейство секреторных белков, характеризуется сходством доменов с N-концевыми доменами 7 цепи ламинина-1 (рис. 7А). Гомологи нетрина также играют роль в росте аксонов у дрозофилы и С. elegans.
Далее аксоны комиссурального нейрона пересекают вентральную среднюю линию, что они делают только однажды, таким образом оставаясь на контралатеральной стороне (рис. 7В, С). Это пересечение облегчается взаимодействиями между двумя адгезивными молекулами клеточной поверхности: TAG-1, которая экспрессируется на поверхности аксона спаечного нейрона, и NrCAM, экспрессируемого на клетках базальной пластинки. После пересечения средней линии экспрессия TAG-1 в аксонах ингибируется сигналами от клеток базальной пластинки, под действием которых в аксонах начинает синтезироваться протеин, называемый robo. Robo представляет собой рецептор для другого белка, называемого slit, который высвобождается клетками базальной пластинки. Взаимодействие между slit и robo отталкивает конусы роста комиссуральных интернейронов. Потеря аттракции к TAG-1—NrCAM контакту и приобретение коротко-дистантного отталкивающего взаимодействия slit-robo предупреждают возможное повторное пересечение аксоном средней линии.
| Рис. 8. Роль нетрина и его рецепторов в аттракции и отталкивании на большом расстоянии. (А) N-концевая аминокислотная часть секреторного белка нетрина состоит из доменов VI и V, которые гомологичны доменам аминокислотного конца 7 цепи ламинина. Домен V содержит три ECG-повтора. С-концевой домен этой цепи не имеет гомологии с ламинином. (В) Рецепторы нетрина имеют внеклеточный домен, однократно пересекающий мембрану, и внутриклеточный домен. Внеклеточный домен семейства DCC рецепторов нетрина имеет четыре иммуноглобулиновых домена (Ig) и шесть повторов фибронектина III (FN III). DCC и их гомологи также могут играть роль молекул адгезии. Семейство рецепторов к нетрину UNC-5 имеет два иммуноглобулиновых домена и два домена, гомологичных тромбоспондину 1-го типа (TSP I), которые расположены внеклеточно, а также длинную цитоплазматическую последовательность. (С) Микрофотографии кусочков дорзального спинного мозга эмбрионов крысы (сверху на каждой из панелей), культивированных вместе с кусочками ткани базальнои пластинки (слева), вместе с CDS клетками, секрета рующи ми рекомбинантный нетрин 1 (в центре), и с контрольными COS клетками (справа). Нижняя пластинка и нетрин 1 оба способны вызывать значительный и направленный рост пучков комиссуральных аксонов из дорзаль ной области спинного мозга. | 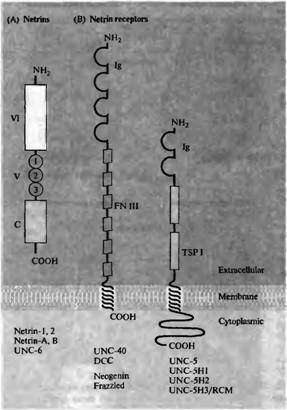 |
Slit и нетрин-1 также диффундируют из базальной пластинки для отталкивания конусов роста мотонейронов. Такие отталкивающие взаимодействия направляют аксоны мотонейронов прочь от спинного мозга в направлении периферии (рис.6D)
Литература:
1. McConnell, S. K. 1995. Constructing the cerebral cortex: Neurogenesis and fate determination. Neuron 15: 761-768.
2. Mueller, В. К. 1999. Growth cone guidance: First steps towards a deeper understanding. Annu. Rev. Neurosci. 22: 351-388.
3. OLeary, D. D., and Wilkinson, D. G. 1999. Eph receptors and heparins in neural development. Curr. Opin. Neurobiol. 9: 65-73.
4. Oppenheim, R.W. 1991. Cell death during development of the nervous system. Annu. Rev. Neurosci. 14: 453-501.