Статья: Генетическая история человечества
Рис. 5. Фрагмент ДНК. Разные индивиды могут отличаться друг от друга по составу нуклеотидов (при постоянной длине фрагмента) или по длине фрагмента (за счёт вставки или выпадения нуклеотидов).
Для исследования используют разные типы ДНК-маркёров. Следует отметить ДНК митохондрий (мтДНК) и Y-хромосомы (Y-ДНК), поскольку они позволяют проследить генетическую историю человечества отдельно по женской и мужской линиям. Первая передаётся только по материнской линии, так как митохондрии находятся в цитоплазме клетки, а цитоплазма потомка (зиготы) определяется цитоплазмой яйцеклетки. Если два человека имеют общего предка женского пола, то различия между их мтДНК покажут, сколько поколений отделяет их от жившей столетия или тысячелетия назад общей прабабушки. Изучение Y-ДНК позволяет проследить эволюционные траектории по отцовской линии, поскольку Y-хромосома передается только по мужской линии. Изучение остальной, аутосомной части генома, которая сосредоточена в хромосомах, наследуется по обеим линиям и в которой представлена большая часть генома человека, также чрезвычайно важно, ибо даёт маркёры для изучения комбинативной изменчивости, одновременно привносимой и с отцовской, и с материнской сторон.
Первое исследование такого рода было проведено с использованием митохондриальной ДНК. Сравнивая данные аборигенов всех континентов, учёные обнаружили, что разнообразие мтДНК выше всего в Южной Африке. Более того, там были обнаружены такие типы, которые нигде больше не встречались. Они оценивались по нуклеотидному составу как наиболее древние. Митохондриальные ДНК населения других континентов были менее разнообразны, их сравнение с мтДНК аборигенов Южной Африки показало, что они возникли как мутационные изменения африканских типов в ходе распространения человечества за пределы Африки.
Изучение географического распределения типов ДНК-маркёров и анализ их мутационных взаимосвязей позволили доказать справедливость монофилетической гипотезы (рис. 4). Более того, их изучение помогает реконструировать миграционные события не только далёких тысячелетий, но и последних веков. Например, эпоха великих географических открытий ХIV-XVI вв. способствовала развитию контактов с неведомыми ранее жителями отдалённых земель. На кораблях были только мужчины, и сейчас генетические исследования аборигенных народов Африки, Океании и Америки показывают присутствие в их ДНК значительной доли типов Y-хромосомы, характерных для европейцев.
«Митохондриальная Ева» и «Y-хромосомный Адам»
По степени разнообразия ДНК-маркёров можно установить, от какого предкового типа и когда они произошли. Более того, поскольку различные типы мтДНК не рекомбинируют, т. е. не обмениваются между собой фрагментами в процессе образования половых клеток, то вероятностный анализ показывает, что все ныне существующие типы мтДНК сводятся в далёком прошлом к одному-единственному прототипу. Почему это происходит? Допустим, существовала небольшая прапопуляция. Если одни женщины в ней имели больше детей, чем другие, то в следующем поколении именно их мтДНК будет встречаться чаще. Их дочери и внучки тоже имели детей, унаследовавшх мтДНК своих предков. При этом случайным образом возникают мутации. Так развивается стохастический процесс в передаче типов мтДНК, в результате чего из поколения в поколение одни типы мтДНК увеличивают свою представительность в популяции, другие — уменьшают.
Согласно теории стохастических процессов, когда-нибудь в популяции останутся потомки только одной мтДНК — от некой единственной женщины, образно именуемой „митохондриальной Евой“. Но и её соплеменницы внесли свой вклад в генофонд отдалённых потомков, поскольку наш геном содержит не только митохондриальную ДНК. Длина мтДНК лишь около 16,5 тыс. нуклеотидов, что ничтожно мало по сравнению с 3 млрд. нуклеотидов ДНК хромосом, основная часть которых представлена 22 аутосомами и X-хромосомой, где комбинируется наследственность по обеим линиям, женской и мужской. Из-за многочисленных мутаций мтДНК ныне живущих людей отличается от данных „Евы“, возраст которой (т. е. точки соединения всех типов мтДНК в прошлом) примерно 200 тысяч лет. Аналогичная ситуация и с Y-хромосомой. Большая часть её не рекомбинирует, и изменчивость ДНК в ней подчиняется тем же стохастическим процессам, что и мтДНК. При этом говорят о „Y-хромосомном Адаме“. Он оказался гораздо моложе „мтЕвы“. Объяснение тому — меньшая, т. н. эффективная численность мужчин на протяжении истории человечества из-за их большей смертности и нередкого отстранения от процесса размножения.
Неандертальский человек — наш прародитель или двоюродный дядя? Дополнительные сведения о нашей родословной были получены при сравнении митохондриальных ДНК современного человека и неандертальца. Более ста лет, с тех пор как в Германии впервые были найдены останки нашего древнего родственника, шли дискуссии о том, кем он нам приходится. Исходя из особенностей строения скелета неандертальца и его географической распространённости, одни учёные считали его нашим прародителем, т. е. линией Homo sapiens, развитие которой привело к человеку современного анатомического типа. Другие детали позволяли считать его независимой эволюционной ветвью, подвидом Homo sapiens, имеющим общего с нами предка, т. е как бы двоюродным дядей. Эти два подвида получили зоологическое название Homo sapiens neanderthalensis и Homo sapiens sapiens.
Учёным удалось прочитать часть вариабельного (контрольного) участка митохондриальной ДНК двух неандертальцев. Первый был найден в Фельдховеровской пещере в Германии, чуть позже был прочитан генетический текст мтДНК неандертальца-ребёнка, обнаруженного на Северном Кавказе в Мезмайской пещере. При сравнении наиболее изменчивых фрагментов мтДНК современного человека и неандертальца были найдены существенные различия: они отличались друг от друга в среднем по 27 нуклеотидным позициям из 370 исследованных. Если сравнить митохондриальные ДНК двух современных людей, то средняя разница будет равна лишь 8 нуклеотидам. Эти расчёты показывают, что наш общий с неандертальцем предок жил примерно 500–700 тыс. лет назад.
Анализ ДНК позволил заключить, что обмен генами между человеком и неандертальцем не происходил или был ничтожно мал. Вероятнее всего, это совершенно отдельные, параллельные эволюционные ветви, произошедшие от общего предка. Хотя для окончательного вывода нужны дополнительные исследования ДНК. Так что неандерталец нам, похоже, эволюционный „дядя“. Приблизительно 300–400 тыс. лет назад произошло окончательное разделение двух ветвей. Неандертальцы первыми расселились по Европе и Азии, затем туда пришли люди современного типа (т.н. кроманьонский человек), и они довольно долго сосуществовали на одной территории. Но около 30 тыс. лет назад неандертальский человек исчез, никаких его следов в более поздних археологических слоях найдено не было. Возможно, он не выдержал конкуренции и был вытеснен и истреблён своим более умным и хитрым родственником, а может быть, существовали и иные причины гибели неандертальцев.
Эволюция популяций и рас
|
Рис. 6. Эволюционное древо популяций человека, по данным о ДНК-маркёрах. Видно, что первично развитие и отделение друг от друга популяций началось в Африке в среднем около 100 тыс. лет назад. Затем одна ветвь вышла из Африки и стала делиться на континентальные ветви. Стрелками указано минимальное время, прошедшее между отделением эволюционных ветвей. Следует иметь в виду, что отделение ветви не означает ещё физического присутствия популяций в этом регионе. Например, ветвь, ведущая от азиатских популяций к американским индейцам, показывает, когда эта ветвь отделилась, но нужно было ещё время, за которое отделившиеся группы достигнут Берингии и перейдут пролив (по: Zhivotovsky et al., 2003). |
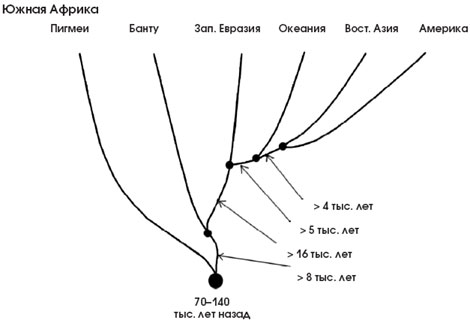
Изучая разнообразие ДНК современных народов, можно оценить численность той прапопуляции, от которой, согласно гипотезе африканского происхождения, произошло всё человечество. Она была невелика — порядка нескольких тысяч. Сопоставляя ДНК-маркёры аборигенов Южной Африки, можно сказать, что примерно 70–150 тыс. лет назад началась интенсивная дифференциация и сложные демографические процессы, сопровождающиеся возникновением разнообразных популяций в пределах Африки. Затем, 50–100 тыс. лет назад, волны переселенцев стали выплёскиваться за пределы Африки и растекаться по другим континентам, что отразилось на своеобразной структуре ДНК-древа (рис. 6).
Исследуя современное население Европы, Азии, Океании, Северной и Южной Америки и зная особенности и скорость мутирования в изучаемых ДНК-маркёрах, можно с определённой степенью точности проследить пути и время миграций людей из Африки. Удивительно то, что генетические данные подтверждаются археологическими находками. Например, структура ДНК свидетельствует о том, что человек появился в Австралии и Новой Гвинее 50–60 тыс. лет назад. Анализ состава химических элементов артефактов указывает на тот же период. В Центральную и Юго-Восточную Азию люди пришли примерно 70 тыс. лет назад, заселение Европы произошло позже, 35–40 тыс. лет назад. Время освоения Америки до сих пор не определено, известно лишь, что люди появились там гораздо позже, чем на других континентах, — от 15 до 35 тыс. лет назад.
|
Рис. 7. Подразделение этнических группировок по географическим районам, осуществлённое по ДНК-маркёрам. Каждая точка представляет собой выборку индивидов определённой этнической группы из данного географического региона, охарактеризованным по четыремстам аутосомным ДНК-маркёрам. Левый и правый рисунки представляют собой разные проекции многомерного пространства различий между выборками. В каждом из регионов прослеживается группировка этнических групп общего происхождения, однако различия между ними уже не столь яркие, как между регионами (по: Zhivotovsky et al., 2003). |
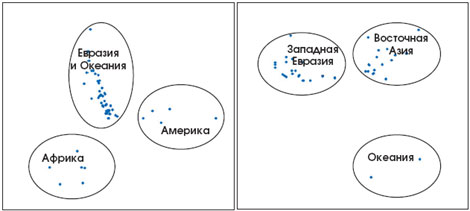
Как возникли современные расы человека и отличаются ли они друг от друга по ДНК? В течение десятков тысяч лет шли процессы миграций и адаптации человека к местным условиям. Допустим, группа людей пришла в Юго-Восточную Азию и осела там на много поколений. Потом часть мигрировала дальше, образуя новую локальную популяцию, которая, однако, имеет общую историю и общих предков с родительской группой, а потому их ДНК более сходны между собой, чем с жителями других континентов. Действительно, население разных материков эволюционно гораздо дальше от общей предковой группы, чем соседние популяции, близкие по родственным связям и демографической истории. За то время, что прошло со времени отделения от общих прародителей, их ДНК стали отличаться друг от друга за счёт накапливающихся в чреде поколений мутаций. Генетические различия между людьми с разных материков мы называем сегодня расовыми признаками. Изучая десятки и сотни ДНК-маркёров, можно почти стопроцентно идентифицировать расу (рис. 7). Чтобы достоверно определить этническую принадлежность индивида в пределах расы и крупного географического региона, потребуются тысячи ДНК-маркёров. А в зонах контакта разных рас и этнических групп это сделать практически невозможно из-за смешения генофондов.
Но генетически мы все вышли из одного гнезда, причём сравнительно недавно в масштабах эволюции (рис. 6).
Дальнейшее развитие рас шло независимо друг от друга: люди адаптировались к климато-географическим условиям, типу питания и ландшафта, складывались язык и культура. Но на формирование народов влияли не только процессы разделения популяций. Новые этносы могли образоваться при смешении групп разной расовой и языковой принадлежности. При этом возникала генетически разнородная этническая общность с единым типом культуры и общим языком. Поэтому сейчас всё большую актуальность приобретают исследования, связанные с изучением генофонда, т. е. всего разнообразия ДНК в популяциях, генетической истории населения отдельных регионов, расово-этнических групп, родословной современных этносов.
Контакт этносов
Особый интерес с этой точки зрения представляет Волго-Уральский регион — в силу особенностей этнической истории населяющих его народов. Здесь встретились две волны расселения, две расы: европеоидная и монголоидная. Следы этого события хранит ДНК проживающих здесь народов (рис. 8). Исследования митохондриальной ДНК и Y-хромосомы позволили рассчитать время формирования народов, заселивших впоследствии данный регион. Это произошло примерно 40–50 тыс. лет назад, что соответствует времени появления современного человека на европейском континенте в эпоху верхнего палеолита.
|
Рис. 8. Сеть мтДНК народов Волго-Уральского региона. Цветом обозначены различные этнические группы, тёмными буквами — разные типы мтДНК, размер кружка — число индивидов с данным типом мтДНК, красными цифрами — мутации. Видно значительное перекрывание изменчивости: в различных этнических группах встречаются одинаковые типы мтДНК, в т. ч. европеоидные, напр. Н., и монголоидные, напр. U. (из: Бермишева и др., 2002). |
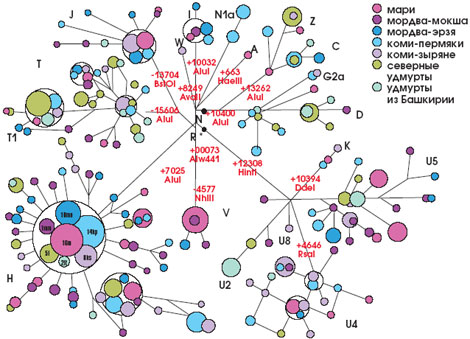
Сравнительный анализ мтДНК 18 народностей Евразии, относящихся к тюркской ветви алтайской языковой семьи, позволил установить западно-восточный градиент увеличения частоты азиатских типов мтДНК на пространстве 8 тыс. км: от 1% у гагаузов из Молдавии до 95–99% у якутов и долган. Соответственно, европеоидные черты наиболее свойственны жителям Западной Евразии, а наименее — населению Восточной Сибири. Народы Волго-Уральского региона, а также узбеки и казахи, т. е. те, кто живёт на границе между Европой и Азией, занимают промежуточное положение. Изучение аутосомных ДНК-маркёров показало наличие в генофонде народов Волго-Уральского региона значительной доли европеоидных черт — от 50 до 90%. Таким образом, оказавшись на границе между Европой и Азией, эти народы сохранили следы смешения двух рас, пришедших одна — с Востока, а другая — с Запада. Кроме того, оказалось, что сходство языков играет меньшую роль, чем географическая близость популяций. Если, например, русские из Рязанской и Курской областей имеют только 2–3% монголоидных типов мтДНК, то русские, проживающие на границе Европы и Азии, имеют их уже 10–12%. Это объясняется их смешением с тюркоязычными народами на территории Волго-Уральского региона.
Таким образом, ДНК какого-либо индивида не даёт возможности определить его национальность, но позволяет выяснить, какого типа у него мтДНК или Y-хромосома: скажем, монголоидной или европеоидной линии. У русских Волго-Уральского региона есть ДНК-маркёры, которые характерны и для марийцев, и для мордвы, и для чувашей, и для башкир, и для татар, и для удмуртов; соответственно, марийцы имеют ДНК-маркёры, встречающиеся у других этнических групп Волго-Уральского региона, и т. д. Эти результаты показывают глубокую генетическую общность давно живущих рядом народов, хотя говорят они на непохожих языках, веруют в разных богов и отличаются культурными традициями.
Все люди — генетические кузены и кузины
Мы все генетически чрезвычайно схожи. Нас отличает друг от друга ряд признаков, которым мы склонны придавать чересчур большое значение (рост, цвет кожи, форма головы и др.), но как они ничтожны в сравнении с нашим почти стопроцентным (99,9%) генетическим сходством! Мы сформировались под влиянием не только генов, но и окружающих нас людей и явлений, причём в становлении личности среда играет неизмеримо большую роль, чем наследственные особенности. Мы все — одна большая генетическая семья, живущая на общей планете. И все распри между людьми возникают на бытовой почве: из-за несоблюдения элементарных норм общежития народов, неуважения к ценностям, особенностям и кажущимся странностям друг друга.
Но представьте себя на необитаемом острове, куда случай забросил ещё одного человека — совсем другой расы, другого вероисповедания, со своим языком и привычками, — 99,9% генетического сходства вас тут же объединят.