Статья: Протеомика
Существование огромного количества разнообразных белков привело к необходимости создания информационных массивов – баз (или банков) данных, в которые заносились бы все известные о них сведения. В настоящее время существует множество общих и специализированных баз данных, которые доступны в Интернете каждому желающему. В общих базах содержатся сведения о всех известных белках живых организмов, т.е. о глобальном протеоме всего живого. Примером такой базы является SwissProt-TrEMBL (Швейцария–Германия), в которой на сегодняшний день содержатся структуры почти 200 000 белков, установленные аналитическими методами, и еще почти 2 млн структур, которые определены в результате трансляции с нуклеотидных последовательностей [10]. На рис. 8 и 9 показано количество существующих белков, которые известны для каждого заданного числа аминокислотных остатков. Оси абсцисс на этих графиках ограничены 2000 остатков, но, как уже сказано выше, хотя и не часто, но встречаются и существенно более крупные молекулы. Из данных, представленных на рисунках, следует, что наибольшее число белков содержит по несколько сотен аминокислотных остатков. К ним относятся ферменты и другие достаточно мобильные молекулы. Среди более крупных белков много таких, которые выполняют опорную или защитную функции, скрепляя биологические структуры и придавая им прочность.
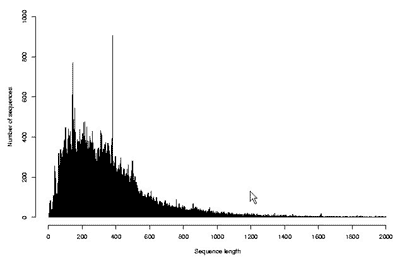
Рис. 8. Распределение известных (выделенных) белков по числу аминокислотных остатков
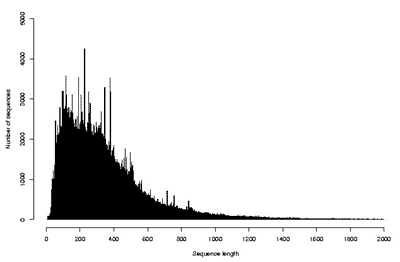
Рис. 9. Распределение транслированных аминокислотных последовательностей по числу минокислотных остатков
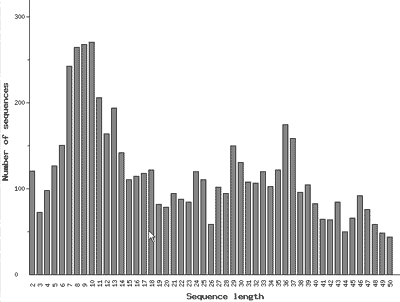
Рис. 10. Распределение известных природных олигопептидов по числу аминокислотных остатков
В глобальном протеоме особое место занимают небольшие очень подвижные молекулы, содержащие не более 50 аминокислотных остатков и обладающие специфическим спектром функциональной активности. Они называются олигопептидами, или просто пептидами. Для них, т.е. для глобального пептидома, создан особый банк данных, который называется EROP-Moscow. Это название представляет собой аббревиатуру от термина Endogenous Regulatory OligoPeptides (эндогенные регуляторные олигопептиды), и указывает на то, что банк создан и базируется в столице нашей страны [11]. На сегодняшний день расшифрована структура почти 6000 олигопептидов, выделенных из представителей всех царств живого. Так же как и крупные белки, количество олигопептидов с заданным числом аминокислотных остатков можно изобразить графически (рис. 10). Судя по графику, чаще всего встречаются олигопептиды, содержащие примерно 8–10 аминокислотных остатков. Среди них в основном содержатся молекулы, которые участвуют в регуляции нервной системы, и поэтому называются нейропептидами. Очевидно, что самые быстрые процессы в живом организме осуществляются с участием нервной системы, поэтому пептидные регуляторы должны быть мобильными и следовательно небольшими. Однако, следует отметить, что, ввиду огромного структурного и функционального разнообразия как белков, так и пептидов, для них до сих пор не создано строгой классификации.
Таким образом, в данном случае задачами биоинформатики являются накопление информации о физико-химических и биологических свойствах белков, анализ этой информации, каталогизация и подготовка информационной базы и вычислительных средств для выявления механизмов их функционирования.
Функциональная протеомика
Наличие в организме того или иного белка дает основание предполагать, что он обладает (или обладал) определенной функцией, а весь протеом служит для того, чтобы осуществлялась полноценная жизнедеятельность всего организма. Функциональная протеомика занимается определением функциональных свойств протеома, и решаемые ею задачи существенно сложнее, чем, например, определение белково-пептидных структур.
Очевидно, что функционирование протеома осуществляется в многокомпонентной среде, в которой присутствует множество молекул других химических классов – сахаров, липидов, простагландинов, различных ионов и многих других, включая молекулы воды. Не исключено, что через некоторое время появятся такие термины, как «сахаром», «липидом» и им подобные. Белковые молекулы взаимодействуют с окружающими их другими или такими же, как и они, структурами, что в конечном итоге приводит к возникновению функциональных реакций сначала на молекулярном уровне, а затем и на макроскопическом. Уже известно множество таких процессов, в том числе с участием белков. Среди них взаимодействие фермента с субстратом, антигена с антителом, пептидов с рецепторами, токсинов с ионными каналами и т.д. (рецепторы и ионные каналы также являются белковыми образованиями). Для выявления механизмов этих процессов проводятся как экспериментальные исследования индивидуальных участников взаимодействия, так и системные исследования средствами биоинформатики. Рассмотрим несколько примеров таких системных подходов.
На рис. 11 показаны представители протеома (в данном случае пептидома) человека – различные гастрины и холецистокинины, которые локализованы в желудочно-кишечном тракте (при написании аминокислотных последовательностей использован стандартный однобуквенный код, расшифровка которого была дана нами ранее [7]). Функциональными частями молекул этих пептидов являются очень схожие правые области. Однако пептиды обладают прямо противоположными поведенческими свойствами: гастрины вызывают у человека ощущение голода, а холецистокинины – сытости. По-видимому, данное различие обусловлено тем, что в первичной последовательности холецистокининов положение остатка тирозина Y сдвинуто на один шаг по сравнению с гастринами. На том же рисунке приведена первичная структура пептида ционина, полученного из представителя простейших хордовых Ciona intestinalis (рис. 12). Его структура гомологична и гастринам, и холецистокининам и характеризуется двумя остатками тирозина, находящимися в тех же положениях, что и у обоих указанных пептидов. К сожалению, функциональные свойства его не изучены. А при должном экспериментальном исследовании можно было бы ответить на вопрос, какова роль химической структуры в целом и остатков тирозина в частности при проявлении противоположных физиологических эффектов.

Рис. 11. Первичные структуры представителей пептидома человека в сравнении со структурой одного из пептидов оболочечника

Рис. 12. Оболочечник Ciona intestinalis, обитающий в Северном море
Другой пример: на рис. 13 приведены аминокислотные последовательности очень похожих молекул, которые также объединены в структурно-гомологичное семейство. Эти молекулы обнаружены у весьма эволюционно далеких живых организмов – от насекомых до млекопитающих. В первой строке дана первичная структура брадикинина, содержащего 9 аминокислотных остатков и встречающегося у многих высших организмов, в том числе и у человека. В течение многих лет химики синтезировали различные неприродные аналоги этой молекулы, чтобы ответить на вопрос, какой ее участок ответственен за взаимодействие с рецептором. Около 30 лет назад были даже синтезированы все возможные фрагменты брадикинина – 8 дипептидов, 7 трипептидов и т.д. (всего возможны 36 фрагментов), величину активности которых затем испытывали в одном и том же биологическом тесте. Результат оказался тривиальным: выяснилось, что максимальную активность проявляет лишь вся молекула целиком, а каждый фрагмент по отдельности обладает либо следовой активностью, либо нулевой. Эту трудоемкую работу не пришлось бы делать, если бы в то время были известны другие брадикинины, приведенные на рис. 13, и средствами биоинформатики они были бы выделены из глобального протеома. Представленное структурно-гомологичное семейство наглядно демонстрирует, что у всех молекул есть область, которая в результате биологической эволюции практически не изменялась (квазиконсервативная область), и она представляет собой молекулу брадикинина высших живых организмов, отобранную как наиболее совершенную в результате эволюционного процесса. Данный пример демонстрирует, что протеомика вместе с биоинформатикой позволяет быстро (и дешево) решать принципиальные научные проблемы.

Рис. 13. Первичные структуры природных пептидов брадикининов, полученных из разных живых организмов. Жирным шрифтом указаны квазиконсервативные области

Рис. 14. Первичные структуры структурно-гомологичного семейства эндотелинов / токсинов
И, наконец, третий пример – структурно-гомологичное семейство эндотелинов млекопитающих и токсинов змей (рис. 14). Несмотря на поразительное сходство структур, их функциональные свойства разительно отличаются друг от друга: одни являются очень полезными регуляторами сосудистого сокращения, а другие – смертельно опасны для жизни. В данном случае мы сталкиваемся с ситуацией, когда первичная структура не несет достаточной информации, способной объяснить причину различия функций, и необходимо более детальное рассмотрение пространственной (третичной) структуры. На рис. 15 и 16 показаны пространственные структуры двух представителей этого семейства – эндотелина-1 и сарафотоксина 6b, полученные с помощью ЯМР-спектроскопии. На рисунках они повернуты так, чтобы достичь максимальной пространственной гомологии. Но полной гомологии не удается получить ни при каком повороте. Следовательно, несмотря на большое сходство первичных структур, взаимодействие их осуществляется с разными рецепторными структурами, а потому и приводит к разным физиологическим эффектам.
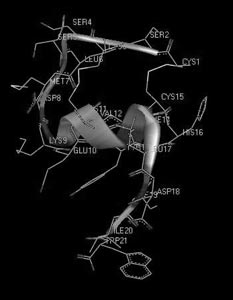
Рис. 15. Пространственная структура сосудосокращающего пептида эндотелина-1 человека
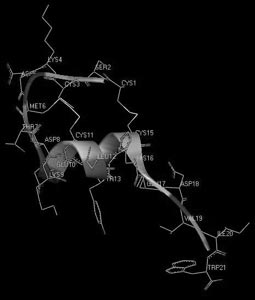
Рис. 16. Пространственная структура сарафотоксина 6b израильской змеи Atractaspis engaddesis
Конечно столь частными примерами невозможно полностью охарактеризовать многообразие функциональной протеомики. Создание представлений об огромной сети взаимодействий белковых и других молекул в организме требует огромного труда и применения всех средств современной биоинформатики. По существу, создание таких представлений еще только начинается. Однако есть основание полагать, что с каждым годом наши познания в этой области будут быстро расти.
Рис. 17. Общие контуры карты метаболизма карбоновых кислот [12]