Учебное пособие: Инфракрасная спектроскопия и спектроскопия кругового дихроизма. Методы определения вторичной структуры белков
Метод "эталонных спектров" [2,3]. М етоды предсказания вторичной структуры белков по их спектрам КД основаны на предположении о том, что спектры КД различных структурных форм, составляющих белковую молекулу, дают аддитивный вклад в спектр КД белка в целом. Это можно записать в следующем виде:
![]() (1.2.1)
(1.2.1)
где ![]() - спектр КД белка (зависимость эллиптичности от длины волны света),
- спектр КД белка (зависимость эллиптичности от длины волны света), ![]() - идеализированный "эталонный спектр" - спектр КД, соответствующий i -ой структурной форме, участвующей в образовании вторичной структуры белка,
- идеализированный "эталонный спектр" - спектр КД, соответствующий i -ой структурной форме, участвующей в образовании вторичной структуры белка, ![]() - доля этой формы во вторичной структуре, причем
- доля этой формы во вторичной структуре, причем
![]() и
и![]() . (1.2.2)
. (1.2.2)
Эталонные спектры ![]() для всех структурных форм могут быть вычислены на основании набора базисных спектров КД (спектров белков с известной вторичной структурой - коэффициентами
для всех структурных форм могут быть вычислены на основании набора базисных спектров КД (спектров белков с известной вторичной структурой - коэффициентами ![]() ) с помощью метода наименьших квадратов и формулы (1.2.1), примененной к каждому базисному спектру. После этого экспериментальный спектр КД исследуемого белка с помощью того же метода наименьших квадратов может быть аппроксимирован по формуле (1.2.1) с использованием вычисленных эталонных спектров. При этом вклад каждого из эталонных спектров будет равен доле соответствующей ему структурной форме в общей структуре белка Такой подход к анализу спектров КД белков был впервые использован в работе [2]. Ниже будет более подробно рассмотрена модификация этого метода [3].
) с помощью метода наименьших квадратов и формулы (1.2.1), примененной к каждому базисному спектру. После этого экспериментальный спектр КД исследуемого белка с помощью того же метода наименьших квадратов может быть аппроксимирован по формуле (1.2.1) с использованием вычисленных эталонных спектров. При этом вклад каждого из эталонных спектров будет равен доле соответствующей ему структурной форме в общей структуре белка Такой подход к анализу спектров КД белков был впервые использован в работе [2]. Ниже будет более подробно рассмотрена модификация этого метода [3].
Принимая в рассмотрение в качестве структурных классов a-спираль (H), b-структуру (b), b-изгиб (t) и “неупорядоченную” форму (R), можем написать:
![]() . (1.2.3)
. (1.2.3)
Суммируя экспериментальные данные, вместо ![]() в уравнение (1.2.3) вводят величину
в уравнение (1.2.3) вводят величину ![]() , учитывающую зависимость эталонного спектра, соответствующего a-спирали, от числа аминокислотных остатков, образующих ее:
, учитывающую зависимость эталонного спектра, соответствующего a-спирали, от числа аминокислотных остатков, образующих ее:
![]() , (1.2.4)
, (1.2.4)
где ![]() и
и ![]() - эталонные спектры для a-спирали из n аминокислотных остатков и для a-спирали “бесконечной” длины, а k - так называемый фактор длины цепи (
- эталонные спектры для a-спирали из n аминокислотных остатков и для a-спирали “бесконечной” длины, а k - так называемый фактор длины цепи (![]() ). Согласно теоретическим расчетам оптической активности a-спирали и экспериментальным данным, спектр КД a-спирали
). Согласно теоретическим расчетам оптической активности a-спирали и экспериментальным данным, спектр КД a-спирали ![]() в диапазоне 185-240 нм может быть разложен на три независимых оптически активных составляющих (n®p*, p®p|| *, p®p^ *), которые можно описать гауссовскими зависимостями:
в диапазоне 185-240 нм может быть разложен на три независимых оптически активных составляющих (n®p*, p®p|| *, p®p^ *), которые можно описать гауссовскими зависимостями:
 , (1.2.5)
, (1.2.5)
где ![]() и
и ![]() - положение максимума и полуширина j -ой гауссовской линии в спектре КД a-спирали, а
- положение максимума и полуширина j -ой гауссовской линии в спектре КД a-спирали, а ![]() - максимальное значение эллиптичности “бесконечной” a-спирали на длине волны
- максимальное значение эллиптичности “бесконечной” a-спирали на длине волны ![]() . В окончательном виде для спектра КД белка можно написать следующее выражение:
. В окончательном виде для спектра КД белка можно написать следующее выражение:
![]() , (1.2.6)
, (1.2.6)
где
 . (1.2.7)
. (1.2.7)
Здесь ![]() - среднее число аминокислот на a-спиральный участок цепи в молекуле белка.
- среднее число аминокислот на a-спиральный участок цепи в молекуле белка.
Параметры ![]() ,
, ![]() ,
, ![]() и
и ![]() в уравнении (1.2.7) были найдены на основе спектра КД миоглобина. Они имеют следующие значения:
в уравнении (1.2.7) были найдены на основе спектра КД миоглобина. Они имеют следующие значения:
| j | ||||
| 1 | 223.4 | 10.8 | -3.73×10 | 2.50 |
| 2 | 206.6 | 8.9 | -3.72×10 | 3.50 |
| 3 | 193.5 | 8.4 | +10.1×10 | 2.50 |
Эти параметры для глобулярных белков с достаточно большой точностью можно считать постоянными. Попытки оценить ![]() для конкретных белков по их спектрам КД оказались ненадежными. Для большинства исследованных белков этот параметр оказался равным примерно 10-11 аминокислотам на a-спиральный сегмент. Распространяя этот факт на все анализируемые белки, авторы данного метода положили
для конкретных белков по их спектрам КД оказались ненадежными. Для большинства исследованных белков этот параметр оказался равным примерно 10-11 аминокислотам на a-спиральный сегмент. Распространяя этот факт на все анализируемые белки, авторы данного метода положили ![]() равным 10.
равным 10.
Вклад b-структуры в спектр КД белка оказывается зависящим от гораздо большего числа параметров: не только от числа аминокислотных остатков на сегмент, но и от числа нитей в данном участке структуры и их направленности, поэтому его описание простым уравнением, подобным уравнению (1.2.7), невозможно. То же самое касается b-изгиба и, особенно, “неупорядоченной” формы, под которой подразумевается все, не относящееся к другим классам. Используемые в данном методе эталонные спектры b-структуры, b-изгиба и “неупорядоченной” формы являются статистически усредненными по белкам, используемым в качестве базисных.
Процедура анализа спектра КД исследуемого белка подразделяется на два этапа. Первый этап заключается в вычислении эталонных спектров структурных элементов, то есть значений ![]() ,
, ![]() ,
, ![]() и
и ![]() для длин волн в диапазоне 185-240 нм с интервалом в 1 нм, на основе экспериментальных спектров КД пятнадцати эталонных белков со значениями
для длин волн в диапазоне 185-240 нм с интервалом в 1 нм, на основе экспериментальных спектров КД пятнадцати эталонных белков со значениями ![]() ,
, ![]() ,
, ![]() ,
, ![]() ,
, ![]() , известными из рентгеноструктурного анализа. Эталонный спектр, соответствующий a-спирали, может быть вычислен непосредственно по формуле (1.2.7). Остальные эталонные спектры находятся из уравнения (1.2.6) с помощью метода наименьших квадратов, причем для уменьшения числа неизвестных в этом уравнении из экспериментального спектра КД каждого эталонного белка исключается вклад a-спиральной формы, вычисленный по формуле (1.2.7). Эталонные спектры, вычисленные с помощью данного метода показаны на рисунке 1.2.1.
, известными из рентгеноструктурного анализа. Эталонный спектр, соответствующий a-спирали, может быть вычислен непосредственно по формуле (1.2.7). Остальные эталонные спектры находятся из уравнения (1.2.6) с помощью метода наименьших квадратов, причем для уменьшения числа неизвестных в этом уравнении из экспериментального спектра КД каждого эталонного белка исключается вклад a-спиральной формы, вычисленный по формуле (1.2.7). Эталонные спектры, вычисленные с помощью данного метода показаны на рисунке 1.2.1.
Когда эталонные спектры найдены, могут быть вычислены коэффициенты ![]() ,
, ![]() ,
, ![]() ,
, ![]() в уравнении (1.2.6), примененном к спектру КД исследуемого белка. Для этого также используется метод наименьших квадратов. Он заключается в подборе таких коэффициентов
в уравнении (1.2.6), примененном к спектру КД исследуемого белка. Для этого также используется метод наименьших квадратов. Он заключается в подборе таких коэффициентов ![]() , что
, что
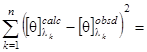 minimum. (1.2.8)
minimum. (1.2.8)
Здесь ![]() - экспериментальный, а
- экспериментальный, а ![]() - рассчитанный по формуле (1.2.6) спектр КД исследуемого белка;
- рассчитанный по формуле (1.2.6) спектр КД исследуемого белка; ![]() - число точек в спектре. Коэффициенты
- число точек в спектре. Коэффициенты ![]() , являющиеся решением уравнения (1.2.8) с учетом условий (1.2.2), представляют собой искомые доли структурных элементов во вторичной структуре белка.
, являющиеся решением уравнения (1.2.8) с учетом условий (1.2.2), представляют собой искомые доли структурных элементов во вторичной структуре белка.
Метод "регуляризации" [4]. Подход к анализу спектра КД белка, лежащий в основе предыдущего метода, заключается в определении эталонных спектров, которые, как можно было бы предполагать, полностью характеризуют структурные элементы, образующие вторичную структуру исследуемого белка. Однако, как показывают экспериментальные данные, ни один эталонный спектр не может точно описать все разновидности таких обширных и достаточно неопределенных классов, как a-спираль, b-структура, b-изгиб и др.
Конформация элементов вторичной структуры глобулярных белков значительно отличается от идеальной. Кроме этого, вклад каждого структурного класса в спектр КД белка зависит от очень многих параметров, о которых упоминалось выше. Для учета всего разнообразия типов вторичной структуры белков требуется расширить исходный набор базисных спектров. В результате возникающей при этом избыточности начальных данных обычный метод наименьших квадратов становится неустойчивым к экспериментальной ошибке и приводит к заведомо неверным результатам. Применение метода "эталонных спектров" в том виде, как он описан в предыдущем пункте, к большому базисному набору спектров оказывается, по сути, некорректным.
Эту проблему частично можно разрешить, заменив метод наименьших квадратов моделью, применение которой, на первый взгляд, не вполне оправдано и адекватно, но зато приводит к устойчивому к экспериментальной ошибке результату даже в случае большого числа параметров. Применение такой стабилизирующей модели позволяет подойти к анализу спектров КД с другой стороны. А именно, появляется возможность прямого представления спктра КД исследуемого белка в виде линейной комбинации базисных спектров. Таким образом удается полностью избежать проблемы, связанной с определением эталонных спектров отдельных структурных классов и проводить более гибкий и точный анализ с использованием реальных белковых спектров.
Рассмотрим данный метод более подробно. Предположим, что нам удалось представить спектр КД исследуемого белка в виде линейной комбинации спектров ![]() базисных белков, структура которых известна из рентгеноструктурного анализа. Обозначим число этих спектров через
базисных белков, структура которых известна из рентгеноструктурного анализа. Обозначим число этих спектров через ![]() (в данном методе
(в данном методе ![]() =16). Тогда можем записать:
=16). Тогда можем записать:
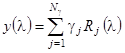 , (1.2.9)
, (1.2.9)
где ![]() - спектр КД (эллиптичность) исследуемого белка.
- спектр КД (эллиптичность) исследуемого белка.