Доклад: Что общего в иммунитете растений и животных
На проходившем в 1975 г. в Ленинграде XII Международном ботаническом конгрессе у американских ученых возник вопрос: “почему секция, посвященная устойчивости растений к болезням, названа “Иммунитет”? Разве русские нашли у растений антитела?” Известный японский фитохимик И.Уритани ответил, что название секции отражает традиции в русской фитопатологической литературе, пришедшие от Н.И.Вавилова, который в 1919 г. написал книгу “Иммунитет растений к инфекционным болезням”. Действительно, в англоязычной фитопатологической литературе термин “иммунитет” встречается редко, чаще пишут “устойчивость” (“resistance”).
Различия в понимании устойчивости к инфекционным болезням у растений и животных зависят от двух причин. Первая связана с факторами иммунитета у тех и других организмов. Известно, что основные успехи медицинской и ветеринарной иммунологии обусловлены использованием антител. Для функционирования этих факторов защиты от инфекции необходимы специализированные клетки и ткани, ответственные за иммунитет, в том числе гуморальная система, способная быстро доставить антитела в любое место. У растений и беспозвоночных животных дифференцировка не столь высока. Каждая клетка растения несет все функции, необходимые для создания целостного организма, в том числе и иммунные. Многочисленные попытки обнаружить антитела у растений были неудачными.
Вторая причина заключается в подходе медика и фитопатолога к защите от болезней. Как писал еще в 1933 г. американский фитопатолог К.Честер, “медик преимущественно занят терапией, фитопатолог - профилактикой”. У медика и даже ветеринара неизмеримо меньше подходов для использования наследственного иммунитета, чем у фитопатолога, поэтому в медицине основные успехи в защите от инфекций лежат на пути иммунизации, т.е. создания приобретенного иммунитета. Фитопатолог же имеет почти неограниченные возможности для искусственного заражения и отбора устойчивых экземпляров среди огромных популяций, а также для гибридизации, инбридинга, мутагенных обработок и проч. Эти преимущества и позволили создать путем селекции устойчивые к болезням сорта многих растений.
Таким образом, болезнеустойчивость позвоночных животных и растений отличается и по биохимическим механизмам, и по методологическим подходам. Это дало основание для шутливого высказывания известного иммунохимика о равновеликом расстоянии между тремя видами иммунитета - животных, растений и дипломатического (сейчас на слуху четвертый - иммунитет депутатский).
Однако по мере углубления знаний о природе болезнеустойчивости, особенно с появлением методов работы с рекомбинантными ДНК, различия в механизмах иммунитета растений и животных перестали казаться столь разительными. Сейчас накапливается все больше данных о сходстве между ними. Об этом и пойдет речь.
Иммуномодуляторы патогенных микроорганизмов
Микроорганизмы, патогенные для животных и растений, выделяют в зараженные ткани и клетки метаболиты (иммуномодуляторы), влияющие на иммунные реакции. Одни из них, индукторы, или элиситоры, хозяин использует для распознавания чужого и индукции в организме защитных реакций. Эти вещества повышают устойчивость как к самим микробам, продуцентам элиситоров, так и к последующему заражению иными патогенными организмами, т.е. представляют собой один из факторов индуцированного приобретенного иммунитета. Другие иммуномодуляторы – иммуносупрессоры, или импедины, – наоборот, подавляют защитные реакции и снижают иммунные свойства. И те, и другие иммуномодуляторы фито- и зоопатогенов сходны по своему строению и механизму воздействия на своих хозяев.
Так, близкие по структуре углеводы (b-глюканы, хитин и др.) служат иммуномодуляторами и растений, и животных. Углеводы фитопатогенов в зависимости от длины цепи, ее конфигурации и генотипа растения могут быть или индукторами, или супрессорами. У животных действие липополисахаридов и пептидоглюканов зависит от их концентрации: малые дозы стимулируют образование антител, большие - подавляют его.
Среди иммуносупрессоров или патотоксинов фитопатогенных грибов много циклических пептидов. Например, пентапептид паразита овса Cochliobolus victoriae (HV-токсин) подавляет активность фермента глициндекарбоксилазы, что приводит к супрессии иммунных ответов, нарушению барьерных свойств мембран, гибели растительных клеток. У человека мутации этого фермента вызывают летальную болезнь - гиперглицинемию. Циклический пептид паразита кукурузы Cochliobolus carbonae (НС-токсин) представляет собой структурный гомолог трапоксина - ингибитора гистонов в клетках животных. Циклоспорин - циклический ундекапептид грибов из рода Tolypoocladium - обладает мощным супрессорным действием на позвоночных животных: подавляет синтез важного межклеточного медиатора (цитокина) интерлейкина-2 в Т-хелперных клетках и активизацию цитотоксических Т-киллеров интерлейкином-2. В связи с этим циклоспорины широко применяют при пересадке органов, ибо они предотвращают иммунное отторжение чужеродной ткани.
Подобные примеры сходства между иммуносупрессорами растений и животных можно найти и среди белков. Так, белок оболочки вируса табачной мозаики (ВТМ, молекулярная масса 17.5 кДа) индуцирует сверхчувствительную реакцию (некроз зараженных клеток) табака; он активен только в виде кристаллических агрегатов. Некроз клеток у животных вызывает фактор некроза опухоли (ФНО, молекулярная масса 17 кДа), продуцируемый Т-лимфоцитами и макрофагами; он активен в виде агрегата из трех молекул и имеет гомологичные последовательности аминокислот со структурным белком сателлита вируса некроза табака. Продукт гена авирулентности avrRxv фитопатогенной бактерии Xanthomonas vesicatoria, поражающей томаты и перцы, индуцирует защитные реакции у устойчивых сортов. Он гомологичен фактору вирулентности yaupj бактерии Yersinia pseudotuberculosum, которая вызывает кишечные инфекции. Транспорт обоих белков через бактериальные мембраны осуществляют специальные белки, имеющие гомологичные области с продуктами hrp-генов у фитопатогенных бактерий и белками секреторной системы типа III у патогенных для человека Yersinia, Shigella и Salmonella.
Не оставляют исключения и липиды. Арахидоновая (эйкозотетраеновая) кислота, продуцируемая возбудителем фитофтороза картофеля Phytophthora infestans, включается в состав фосфолипидов зараженных клеток, окисляется там ферментом липооксигеназой до эйкозаноидов. Эта кислота - активный индуктор окислительного взрыва, синтеза растительных антибиотиков (фенолов, фитоалексинов) и реакции сверхчувствительности. У животных она - предшественник эйкозаноидов, окисляется липо- и циклооксигеназами до лейкотриенов и простагландинов - медиаторов циклических нуклеотидов и агрегации тромбоцитов.
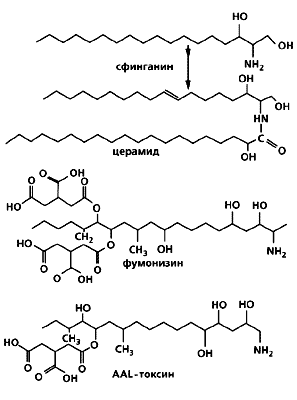 |
Синтез церамида (регулятора клеточного цикла и апоптоза) из сфинганина и структура грибных антиметаболитов фумонизина и AAL-токсина. У растений эти токсины подавляют защитные реакции и транспорт сахаров, приводят к неопластическому росту и некрозам. У человека и сельскохозяйственных животных фумонизин вызывает гепатотоксикозы, разные формы неоплазмозов и гибель клеток. |
Иммунные молекулы
Строение антител - иммуноглобулинов - хорошо известно: две полипептидные цепи - тяжелая (H, молекулярная масса ~50 кДa) и легкая (L, молекулярная масса ~25 кДa), соединенные дисульфидными связями. Каждая цепь содержит чередующиеся константные (С) и вариабельные (V) участки. Огромное разнообразие антител в организме, позволяющее узнавать множество чужеродных антигенов, обусловлено уникальным генетическим контролем их синтеза.
Продукты генов устойчивости растений названы R-белками (от англ. resistance - устойчивость). Структуру первых трех R-белков (из томатов, арабидопсиса и табака) установили сравнительно недавно - в 1993 г. С тех пор их описано несколько десятков.
В состав R-белков входит несколько структур, обеспечивающих, с одной стороны, взаимодействие с лигандом (элиситором) и с молекулами-мишенями (в частности, с ДНК), а с другой - передачу сигнала на другие молекулы - циклические нуклеотиды, протеинкиназы и др. Полипептидные цепи R-белков образованы комбинациями фрагментов, что и обеспечивает растениям из разных классов и порядков устойчивость к вирусам, бактериям, грибам и даже нематодам. Следовательно, эти структуры возникли очень давно, до расхождения предковых растений на современные классы и порядки, выполняли иные, более общие, функции, связанные с рецепцией и передачей сигнала от возбужденного рецептора к генам. Значит, их иммунная функция вторична.
Гены устойчивости растений, контролирующие синтез R-белков, расположены на хромосомах неслучайно. Широко распространены три типа локализации: одиночное диаллельное (один локус с двумя аллелями, контролирующими восприимчивость и устойчивость, которая, как правило, доминантна); одиночное мультиаллельное (один локус с множеством кодоминантных аллелей, контролирующих устойчивость к разным видам и расам патогена); сцепленное (большое число ди- или мультиаллельных локусов сцеплены и образуют блок фенотипически сходных генов, определяющих устойчивость к одной или нескольким болезням). Вот некоторые примеры.
Ген устойчивости риса к бактерии Xanthomonas oryzae Xa21 входит в мультигенное семейство, состоящее по крайней мере из восьми членов, которые картируются в одном локусе на хромосоме 11. На коротком плече
V-й хромосомы ячменя находится блок из пяти локусов, определяющих устойчивость к мучнистой росе; в одном из локусов - Mla - картировано около 20 кодоминантных аллелей. Устойчивость кукурузы к расам возбудителя северной ржавчины Puccinia sorgi контролируют пять локусов. Два из них - Rpl и Rp5 - тесно сцеплены, причем в локусе Rpl картировано 14 аллельных и тесно сцепленных генов. Гены устойчивости томатов к кладоспориозу (возбудитель - гриб Cladosporium fulvum ) образуют несколько групп сцепления. Тесно сцеплены гены Cfl, Cf-4 и Сp, расположенные на коротком плече хромосомы 1, а также Cf-2 и С-5 на коротком плече хромосомы 6. Последние, вероятно, аллельны, так как не рекомбинируют у гибридного потомства.
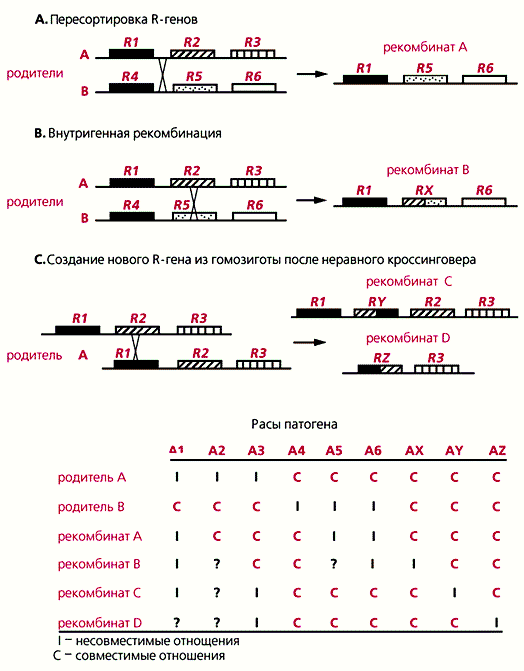
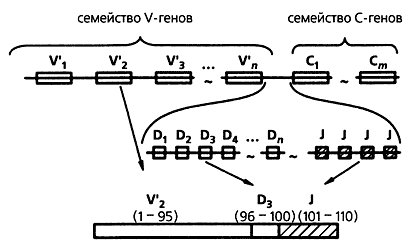
Схема участка хромосомы, включающего семейства вариабельных, V, и константных, C, генов (вверху). Численность V-генов в отдельных семействах варьирует от 50 до 1000, а C-генов - от 1 до 9. Внизу - сборка V-гена: V-фрагмент - контролирующий первые 95 аминокислот, D - следующие за ним от 2 до 10, J - концевой фрагмент V-гена, контролирующий от 5 до 10 концевых аминокислот. Районы ДНК, разделяющие V-D и D-J фрагменты, вырезаются и утрачиваются при сборке гена.
Возникновение кластеров сцепленных генов, функционально и, вероятно, структурно сходных, обусловлено внутри- или межгенными обменами участков ДНК, имеющих прямые (аbсd - а’b’с’d’) или инвертированные (аbсd - d’с’b’a’) повторяющиеся последовательности. Нити ДНК двух родителей в мейозе могут спариваться по гомологичным последовательностям, которые находятся в негомологичных участках хромосом. Это приводит к неравному кроссинговеру. В результате у одного родителя участок хромосомы теряется (делетируется), а у второго - удваивается (дуплицируется), т.е. вместо одного гена появляется два сцепленных.
С этой точки зрения наибольший интерес представляет LRR-область R-белков с многочисленными повторяющимися последовательностями. Благодаря точковым мутациям, делециям, инверсиям такая структура обеспечивает генетическую реорганизацию. Экспериментально показано, что мутации, изменяющие реакцию на заражение авирулентными расами патогенов, картированы в LRR-области.
Большую роль в увеличении генетического разнообразия семейств генов устойчивости могут играть разрывы кодирующей части ДНК, вызванные внедрением мобильных генетических элементов. Так, у риса в локусе белка Ха21 обнаружены транспозоны Truncator и Retrofit, приводящие к разрывам и образованию самостоятельных рамок считывания. Этот белок сочетает свойства двух белков томата - мембранного рецептора и фермента протеинкиназы. Возможно, возникновение генов С/9 и Pto обусловлено разрывом предшественника, подобного гену Ха21.
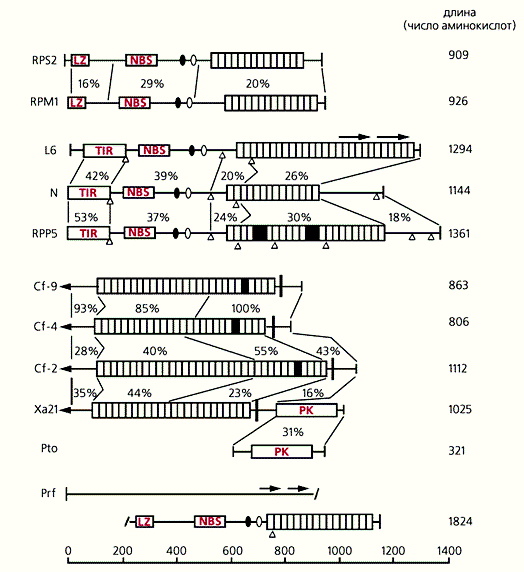
Схема строения R-белков. Заштрихованы С-концевые домены, которые содержат повторяющиеся последовательности, богатые лейцином (LRR-области). Каждый повтор состоит из 23-24 аминокислот. Эта структура, встречающаяся во многих белках эвкариот, осуществляет белковые взаимодействия, т.е. служит рецептором, связывающимся с лигандом - элиситором патогена. NBS (Nucleotide Binding Sites) - сигнальная область, связывающаяся с АТФ и ГТФ, вследствие чего она может активировать киназы или сигнальные G-белки. LZ (Leucine Zipper Region) - область лейциновой “застежки”, которая участвует в формировании спирализованных структур, ответственных за димеризацию или специфическое взаимодействие с другими белками. TIR (Toll/Interleukin-1 Resistance) - область гомологии с цитоплазматическим доменом Toll-белка дрозофилы и рецептором интерлейкина-1 млекопитающих. Возможно, TIR-область растительных R-белков несет сходные функции. РК - серин-треониновая протеинкиназа - фактор активации транскрипции и других сигнальных путей. Слева обозначены гены растений, кодирующие R-белки: RPS и RPM - гены устойчивости арабидопсиса к бактериям из рода Pseudomonas; L - ген устойчивости льна к ржавчине; N - ген устойчивости табака к вирусу табачной мозаики; RPP - ген устойчивости арабидопсиса к ложной мучнистой росе; Cf - гены устойчивости томатов к грибу Cladosporium fulvum; Xa - ген устойчивости риса к бактерии Xanthomonas oryzae; Pto и Prf - система генов, контролирующих устойчивость томатов к бактерии Psiulomonas tomato. Справа - содержание аминокислот в белках. Внутренние цифры - проценты идентичных последовательностей в специфических областях. Стрелки указывают направления транскрипции, а треугольники - положение интронов.
Разные типы рекомбинаций (A, B, C) на хромосомном сегменте, содержащем локусы устойчивости растений к болезням (А – С). Внизу - гипотетические изменения устойчивости к расам патогенного микроорганизма, возникающие в результате рекомбинаций.
Таким образом, аналогично множественным генам иммуноглобулинов у млекопитающих, обеспечивающих синтез антител, у растений имеются белки, которые кодируются семейством сцепленных генов. Высокая вариабельность их генопродуктов позволяет быстро реагировать на заражение новыми вирулентными видами и расами паразитов.
Передача сигнала и иммунный ответ
Между возбуждением рецептора и активацией генов иммунного ответа происходит трансдукция - передача сигнала, в ходе которой он многократно умножается. Процесс осуществляется сигнальными системами (как правило, общими для всех клеток), участвующими в регуляции разных сторон жизнедеятельности организма. Некоторые сигнальные системы функционируют в зараженных и больных клетках и тканях.
--> ЧИТАТЬ ПОЛНОСТЬЮ <--