Контрольная работа: Отдел сине-зеленые водоросли
– Cyanophyta
К данному отделу относят одноклеточные, колониальные и нитчатые водоросли, различно окрашенные в зависимости от соотношения пигментов, представленных кроме хлорофилла а и каротиноидов еще синими пигментами – фикоцианином и аллофикоцианином и красным фикоэритрином. Характерно также полное отсутствие жгутиковых стадий. Половой процесс у сине-зеленых водорослей не наблюдался.
Цитология. При рассматривании клетки сине-зеленых водорослей в оптическом микроскопе можно видеть, что окруженная оболочкой цитоплазма, как правило, лишена вакуолей с клеточным соком, окрашена в периферических частях и бесцветна в центре. Резкой границы между хроматоплазмой и центроплазмой нет. В центроплазме локализирована ДНК, что позволяет рассматривать ее как гомолог клеточного ядра, хотя ядерная оболочка и ядрышки отсутствуют. Кроме того, в цитоплазме находятся включения запасного характера и часто – полости, наполненные смесью газов, по составу близкой к воздуху. У одних видов псевдовакуоли встречаются на протяжении всей вегетативной жизни, у других – только на определенных стадиях развития.
В электронном микроскопе клетки сине-зеленых водорослей имеют следующее строение. Клеточная стенка состоит из четырех четко разграниченных слоев, обозначаемых Lx , L2 , L3 , L4 . Непосредственно кнаружи от цитоплазматической мембраны расположен электронно-прозрачный слойза которым следует электронно-плотный слой, состоящий из муреина – основного компонента клеточной оболочки бактерий. У эукариотических. водорослей и грибов это вещество не обнаружено. Таким образом, и по химизму клеточной оболочки сине-зеленые водоросли, нередко называемые теперь сине-зелеными бактериями, более близки к бактериям, чем к эукариотическим водорослям. Именно этот муреиновый слой L2 определяет прочность стенки. Будучи изолирован, этот слой клеточной стенки способен сохранять форму всей клетки. Следующие за муреиновым слоем слои – электронно-прозрачный L3 и мембраноподобный L4 – образованы углеводами и в отличие от слоя L, гибкие, пластичные.
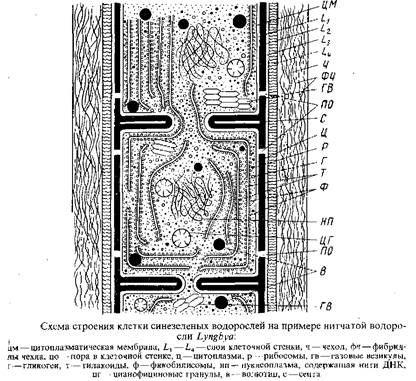
Обусловленная этими слоями пластичность клеточной стенки допускает изменения формы клетки. На уровне слоя L3 у осциллатории были обнаружены непрерывные параллельные ряды, по-видимому, протеиновых микрофибрилл, которые по спирали окружают трихом. Ориентация этих микрофибрилл совпадает с направлением вращения трихома во время его скольжения. Было высказано предположение, что волнообразные изгибы этих рядов микрофибрилл могут обусловливать скользящее движение трихома на твердом субстрате. Упомянутые четыре слоя наблюдаются в продольных стенках нитчатых сине-зеленых водорослей. Поперечные стенки, или септы, нитчатых сине-зеленых водорослей образованы только слоями L1 и L2 . У одноклеточных форм наружные слои L3 и L4 образуются на септах только после начала разъединения клеток. Как в продольных стенках, так и в септах нитчатых сине-зеленых водорослей имеются поры, через которые соединяются цитоплазматические мембраны и протопласты соседних клеток; эти цитоплазматические тяжи называются микроплазмодесмами.
Подсчеты показали, что между двумя вегетативными клетками одного из видов анабенопсиса в септе может быть до 4000 микроплазмодесм.
У многих сине-зеленых водорослей клеточные стенки покрыты слизистым слоем, который может быть толстым и плотным и образует чехлы или капсулы, обычно заключающие несколько клеток, или же слизь представлена в виде тонкого жидкого слоя. Слизь предохраняет клетки от высыхания и, по-видимому, принимает участие в процессе скользящего движения. Тонкая структура слизи – фибриллярная.
Преимущественно в периферической цитоплазме – хроматоплазме – локализованы тилакоиды, способные иногда проникать во все части клетки. Тилакоиды не отграничены от цитоплазмы мембранами, как у других хлорофиллоносных растений, т.е. истинные хлоропласты, одетые оболочкой, здесь отсутствуют. Периферически расположенные тилакоиды обычно ориентированы параллельно продольной клеточной стенке, но иногда они расположены перпендикулярно к продольной стенке. Изредка тилакоиды рассеяны по всей клетке.
Тилакоиды сине-зеленых водорослей никогда не образуют групп, как это свойственно эукариотическим водорослям, за исключением красных, где тилакоиды также расположены одиночно. Добавочные пигменты сине-зеленых водорослей в форме гранул – фикобилисом – локализованы на поверхности тилакоидов.
Центр клетки, обычно свободный от тилакоидов, занят нуклеоплазмой, не отграниченной от остальной цитоплазмы ядерной оболочкой» В нуклеоплазме находятся фибриллы ДНК. Для ядерного материала сине-зеленых водорослей, как и для бактерий и фагов, характерно отсутствие гистонов, чем они отличаются от остальных эукариотических организмов, ядерная субстанция которых содержит помимо ДНК, ядерный белок.
В цитоплазме, не занятой тилакоидами и нуклеоплазмой, находятся рибосомы и запасные вещества: гликоген, волютин, цианофициновые гранулы.
Газовые вакуоли, рассеянные по всей клетке или располагающиеся у поперечных перегородок, состоят из тесно упакованных, наподобие сот, полых, одетых мембраной субъединиц – газовых везикул. Эти везикулы имеют форму полых цилиндрических трубок с коническими шапочками у концов. Они могут быть изолированы из клетки без изменения их формы. Подвергнутые определенному давлению цилиндрические части газовых везикул спадаются, при этом конические шапочки могут отламываться. Химические анализы изолированных газовых везикул показали, что их мембраны отличаются от типичной элементарной мембраны отсутствием липидов. Мембраны газовых везикул состоят только из белков.
Размножение. Перед клеточным делением количество ДНК удваивается, а по мере деления клетки удвоенное количество ДНК разделяется пополам. Клетки сине-зеленых водорослей делятся таким образом, что на боковой стенке их возникает кольцевая складка, образованная цитоплазматической мембраной и внутренними слоями клеточной оболочки. Разрастаясь в центростремительном направлении, yiaскладка смыкается наподобие ирисовой диафрагмы микроскопа, формируя поперечную перегородку – септу, перерезающую тилакоиды и содержимое клетки.
В середине поперечно перерезанной септы в процессе утолщения муреинового слоя сравнительно рано становится различимым слой более низкой плотности – срединная ламелла. У одноклеточных форм и многих ностокальных, у которых образование клеточной перегородки сопровождается появлением более или менее глубокой перетяжки между дочерними клетками, клетки разъединяются путем центрипетального врастания в плоскость срединной ламеллы между слоями муреина (L2 ) наружных слоев клеточной оболочки. Таким образом, у многих ностокальных разрыв трихома при образовании гормогониев происходит между клетками – интерцеллюларно. Совсем иначе разъединяются клетки у многих осциллаториальных и сцитонемовых. Хотя муреиновые слои зрелой септы разъединены срединной ламеллой, расщепления септы и расхождения клеток в этой области не происходит, так как муреиновые слои закреплены у периферического утолщения края диска септы. Слабым местом в муреиновом одеянии клеток у форм с нерасщепляющейся септой, по-видимому, является набор многочисленных мелких соединительных пор, расположенных по окружности по обе стороны от септы – вдоль соединения ее с продольными стенками. Разрыв трихомов при образовании гормогониев происходит вдоль соединительных пор – поперек клеток – трансцеллюларно и связан с гибелью клетки, «приносимой в жертву» – «жертвенной клетки». При этом ненарушенные септы между «жертвенной клеткой» и двумя соседними с ней клетками становятся стенками конечных клеток сформировавшихся гормогониев. Остатки клеточных стенок «жертвенных клеток» могут служить доказательством того, что разрыв произошел не благодаря расщеплению септ между живыми клетками, а трансцеллюларно. Такие остатки оболочек отмерших клеток, по которым происходит разрыв, часто можно видеть в оптическом микроскопе на концах трихомов осциллаторий, Как интерцеллюларным, так и трансцеллюларным разрывам способствуют многократные изгибы трихомов.
У одних нитчатых сине-зеленых водорослей все клетки нити одинаковы – это гомоцитные талломы у других нити состоят из вегетативных клетки гетероцист, нередко наблюдаются и споры – гещероцитные формы. Гетероцисты и споры – это специализированные клетки, образующиеся путём «дифференцировки вегетативных клеток. Один из наиболее заметных признаков гетероцист – их сильно утолщенные стенки. Клеточные стенки гетероцист, как и вегетативных клеток, состоят из слоев Lx –L4 , но кнаружи от них развиваются еще три специальных слоя оболочки гетероцист.

Непосредственно кнаружи от клеточной стенки располагается пластинчатый слой, особенно толстый у глубокой перетяжки между гетероцистой и соседней вегетативной клеткой, где имеется поровый канал – «пора», пересеченная септой, пронизанной микроплазмодесмами, посредством которых сообщаются гетероциста и вегетативная клетка. Кнаружи от пластинчатого слоя находится гомогенный слой, который без резкой границы переходит в самый наружный фибриллярный слой. Благодаря толстой обвертке, образующейся кнаружи от клеточной оболочки, гетероцисты выдерживают механические воздействия, разрушающие более тонкостенные вегетативные клетки.
Другой признак зрелых гетероцист, легко наблюдаемый в оптический микроскоп, – гомогенность клеточного содержимого, которая обусловлена исчезновением гранулярных включений, характерных для вегетативных клеток. Ни волютина, ни цианофициновых зерен в полностью развитых гетероцистах нет. Единственные гранулярные структуры, которые можно различить на электронных микрофотографиях зрелых гетероцист, – рибосомы. Газовые вакуоли, если они были в вегетативных клетках, при дифференцировке их в гетероцисты также исчезают.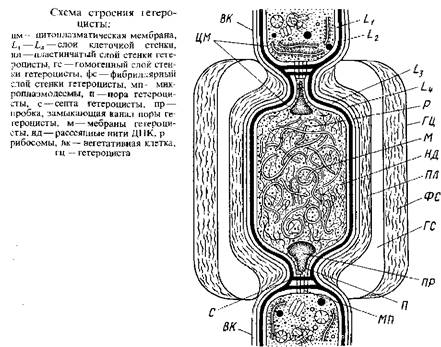
Наконец, дифференцировка гетероцист сопровождается реорганизацией мембранной системы клетки: тилакоиды разрушаются и формируются новые плотно упакованные мембраны. Преобразование мембранной системы сопряжено с изменениями в пигментном составе: в гетероцистах обнаруживаются хлорофилл и каротиноиды, но почти нет фикоцианина, аллофикоцианина и фикоэритрина.
Фибриллы ДНК, в вегетативных клетках обычно сконцентрированные в нуклеоплазме, в гетероцистах рассеяны по всей цитоплазме.
В многочисленных исследованиях было показано, что образование гетероцист, как и процесс фиксации атмосферного азота, ингибируется присутствием в питательной среде связанного азота, особенно аммонийного. Исходя из этих данных стало возможным при выращивании на средах, содержащих связанный, особенно аммонийный азот, искусственно получать трихомы без гетероцист у родов, в норме характеризующихся наличием гетероцист, например у анабен, ностоков и др. При перенесении такого безгетероцистного материала в среду, свободную от связанного азота, можно проследить все стадии развития гетероцист из вегетативных клеток. Дифференцировка гетероцист начинается с образования самого наружного фиброзного слоя ее оболочки кнаружи от клеточной стенки дифференцирующейся вегатативной клетки. Образование фиброзного слоя и следующего гомогенного слоя сопровождается разрушением гранулярных цитоплазматических включений и тилакоидов. На конечной стадии развития гетероцист откладывается самый внутренний пластинчатый слой оболочки гетероцисты, особенно толстый вокруг порового канала, и формируется развитая мембранная система, по-видимому, путем соединения мелких пузырьков, возникающих на местах предшествующих тилакоидов.
Давно известно, что у таких родов, как анабена, носток, характеризующихся интеркалярными гетероцистами, в гетероцисты превращаются не любые вегетативные клетки, а лишь некоторые, расположенные на определенном расстоянии друг от друга. Сравнительно недавно было высказано предположение, что гетероцисты анабены выделяют ингибитор, подавляющий дифференцировку вегетативных клеток в гетероцисты. Последние возникают из вегетативных клеток, расположенных вне ингибирующих зон, окружающих ранее образовавшиеся гетероцисты.
В ряде работ эту гипотезу удалось подтвердить экспериментально. Так, Д. Тивари из дикого штамма Nostoclinckia, способного осуществлять процесс азотфиксации, выделил мутанты, не фиксирующие атмосферный азот, по сохранившие возможность осуществлять развитие, ведущее к дифференцировке гетероцист. На средах, не содержащих азота, у этих штаммов появлялись иптеркалярные гетероцисты. Однако в отличие от дикого исходного штамма вегетативные клетки по соседству с возникшими гетероцистами также превращались в гетероцисты, что приводило к образованию цепочек гетероцист. Напрашивается вывод, что ингибитор – это дериват какого-то непосредственного продукта азотфиксации. Он максимально концентрируется вокруг функционирующих гетероцист, регулируя распределение гетероцист вдоль нити. У других родов гетероцисты занимают терминальное положение в трихоме; бывают и латеральные гетероцисты.
У очень многих гетероцитных форм, в зрелом состоянии характеризующихся интеркалярными гетероцистами, их молодые проростки имеют терминальные гетероцисты.
Относительно функции гетероцист высказывались разные предположения. У гетероцитных форм распад нитей на отдельные участки, из которых развиваются новые талломы, обычно происходит по гетероцистам, соответственно им приписывалась и известная роль в вегетативном размножении. Наблюдаемые иногда случаи деления содержимого гетероцист позволяли предположить их первоначальную роль в качестве клеток, предназначенных для размножения, И действительно, при культивировании в определенных условиях описано довольно много фактов прорастания гетероцист. Так, у Anabaenacylindrica, выращенной в безазотистой среде, при перенесении ее на среду с хлоридом аммония 3–10% гетероцист прорастали. У неспорулирующего мутанта одного из видов Gloeotrichiaпри культивировании в среде с хлоридом аммония прорастало 83% гетероцист. Сравнительно недавно было даже высказано предположение, что исчезновение гетероцист, наблюдаемое на средах, в состав которых входит связанный азот, объясняется как раз их прорастанием. Однако в естественных условиях прорастание гетероцист – редкое явление. Против их роли в качестве клеток бесполого размножения свидетельствует не только то, что они лишены запасных продуктов, которые могли бы поддержать начальные стадии прорастания, но и то, что при отделении их от нити, они, как правило, быстро разрушаются, возможно, оттого, что в отличие от спор не защищены со всех сторон оболочкой. Согласно последним данным в гетероцистах происходит фиксация атмосферного азота в аэробных условиях.
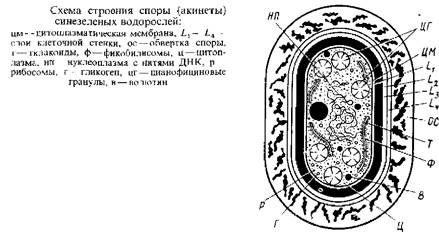
Другими специализированными клетками, которые также образуются путем дифференцировки вегетативных клеток, являются споры – акинеты. Это обычно более крупные, чем вегетативные, клетки, толстостенные, причем споровый покров окружает акинету полностью в отличие от гетероцист, которые через поровый канал сохраняют контакт с соседними вегетативными клетками. При дифференцировке спор из вегетативных клеток заметно утолщается L2 слой и, кроме того, кнаружи от клеточной оболочки образуется широкая обвертка. В отличие от гетероцист зрелые споры переполнены запасными гранулярными включениями, особенно цианофициновыми зернами. Расположение и структура тилакоидов в спорах те же, что и в вегетативных клетках. Содержание ДНК в спорах заметно возрастает, иногда в 20–30 раз по сравнению с содержанием ее в вегетативных клетках. Как и в случае гетероцист, вегетативные клетки, превращающиеся в споры, утрачивают газовые вакуоли.
Спорообразование у сине-зеленых водорослей стимулируется разными факторами среды, но особенно фосфором. Споры могут выдерживать высыхание и другие неблагоприятные условия. Они могут прорастать сразу же после образования, без какого-либо периода покоя, при помещении в благоприятные условия. При отсутствии таких условий споры долго сохраняют жизнеспособность. Например, споры Anabaenaspiroides, A. flos-aquae, Aphanizomenonflos-aquae, выделенные из донных осадков, возраст которых составлял от 18 до 64 лет, хорошо прорастали на минеральной среде в лабораторных условиях. При прорастании споры образуется один проросток, освобождающийся через разрыв оболочки.
--> ЧИТАТЬ ПОЛНОСТЬЮ <--