Контрольная работа: Структура иммуноглобулинов
Содержание
1. Первичная структура Н-и L-цепей иммуноглобулинов
2. Трехмерная структура иммуноглобулинов
3. Антигенсвязывающие центры антител
1. Первичная структура Н-и L-цепей иммуноглобулинов
При исследовании аминокислотной последовательности было обнаружено, что все легкие и тяжелые цепи имеют одну принципиальную структурную особенность: они состоят из двух частей — вариабельной и константной. Вариабельные части легких цепей сильно отличаются друг от друга у всех исследованных иммуноглобулинов, а константные имеют близкую аминокислотную последовательность в пределах одного класса как легких, так и тяжелых цепей.
Постоянная или константная часть легких цепей для каждого из х- или Я-типов цепей включает 107 а. о. СООН-концевого участка. Исключение составляют различия в нескольких аминокислотных остатках, например в положении 191 и 153 в цепях человека или в положениях 190 и 152 в Я-цепях, обусловленные генетическими вариантами.
Оставшаяся последовательность аминокислотных остатков в NHa-концевой половине легких цепей образует так называемую вариабельную область. Частота замен отдельных аминокислот в вариабельных областях легких цепей иммуноглобулинов зависит от положения аминокислотного остатка в полипептидной цепи и максимальна около аминокислот в положениях 30, 50 и 95. Эти участки, получившие название гипервариабельных, принимают непосредственное участие в связывании антигена и входят в состав антигенсвязывающего центра.
Вариабельная область Н-цепей в МН2-концевой части несколько длиннее соответствующей области L-цепей и включает 118—124 а. о. Гипервариабельными участками являются четыре аминокислотные последовательности: 31—37, 51—68, 84—91 и 101—110, также непосредственно контактирующие с антигеном в антигенсвязывающем центре. Последовательности аминокислотных остатков вариабельных областей Н-цепей различных иммуноглобулинов по степени гомологии могут быть разделены на три подгруппы, внутри каждой из которых эта величина составляет 8,0—90%.
Константная область тяжелой цепи приблизительно в три раза длиннее вариабельной.
2. Трехмерная структура иммуноглобулинов
В каждой из легких цепей молекул антител существуют две внутрицепочечные дисульфидные связи, число таких связей в тяжелых цепях различно. Каждый из внутрицепочечных дисульфидных мостиков образует петлю из 55—70 а.о.
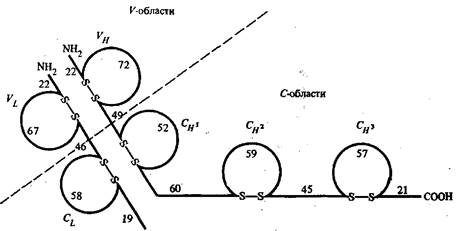
Рис. 1 - Схема расположения внутрицепочечных дисульфидных связей в легких и тяжелых цепях молекулы IgG человека
На рис. 1 приведено схематическое изображение расположения дисульфидных связей вдоль тяжелых и легких цепей IgG человека с указанием среднего числа аминокислотных остатков в составе каждой из петель. По данным рентгеноструктурного анализа, участки пептидных цепей вблизи петли образуют глобулу, в которую включается примерно 110 а. о. Такие глобулы получили название доменов. Н2-концевые домены тяжелых и легких цепей относятся к вариабельным частям, а СООН-концевые домены — к константным частям. Было установлено, что аминокислотные последовательности константных доменов из тяжелых цепей в пределах одного класса являются весьма сходными между собой.
Н2-концевой домен тяжелой цепи обозначают как Ун, а три последующих в константной области тяжелой цепи — как Сн1. Сн2 и СнЗ. А и у тяжелые цепи содержат по 3 константных домена, обозначаемых как Са1, Са2, Са3 для IgA и Cvl, Су2, С£ для IgG, а более длинные ц- и е-цепи — по четыре.
Рентгеноструктурный анализ, выполненный для нескольких белков — димера Я-цепей, двух димеров F-областей Fab-фрагментов IgG и IgA, а также Fc-фрагмента IgG, позволил установить пространственное расположение отдельных участков полипептидных цепей и доменов. Домены L- и Н-цепей представляют собой полусферы, соединенные вдоль одной цепи линейными полипептидиыми отрезками, доступными растворителю и действию протеолитических ферментов. Их основным структурным элементом являются два почти параллельных слоя, один из которых состоит из четырех линейных антипараллельных отрезков цепи, второй — из трех. В состав этих р-слоев входит около 60% антипараллельные сегменты fxl — fx4 и fyl — fy3 обра-ауют два практически параллельных слоя Р-складчатой структуры. Между ними лежат сегменты, образующие изгибы цепи, спирали н т.п.; V- н С-домены отличаются частью сегментов; V-домен имеет добавочную петлю Е; черный прямоугольник соответствует внутрицепочечной дисульфидной связи всех аминокислотных остатков области. Боковые заместители гидрофобных аминокислот образуют плотно упакованное ядро между р-слоями. Дисульфидная связь между р-слоями, являющаяся существенным элементом жесткости структуры доменов, находится в самом гидрофобном ядре и поэтому в водных растворах недоступна для восстанавливающих агентов.
Участков с а-спиральной конформацией в структуре доменов практически не содержится. По сравнению с С-доменами К-обла-сти несколько длиннее и содержат дополнительную петлю Е. В К-областях образующие ядро аминокислотные остатки с гидрофобными боковыми заместителями, а также остатки в изгибах полипептидной цепи, обеспечивающие ее необходимый поворот, являются консервативными или полуинвариантными. Гипервариабельные остатки располагаются на изгибах цепи таким образом, что оказываются пространственно сближенными.
Взаимодействие между Н-цепями в Fab-фрагменте обеспечивается множественными контактами между парами доменов Уъ и. Vn, Cl и Сн1. При ассоциации'Cl и Сн1 домены обращены друг к другу четырехцепочечными р-слоями и взаимодействуют вдоль обширной зоны.свободной от растворителя. В контактах, имеющих в основном гидрофобный характер, участвует около 30 а. о. Наоборот, Vi, и Ун-домены ориентированы друг к другу трехце-почечными р-слоями. Основную роль в стабилизации образующейся структуры играют взаимодействия между консервативными и полуинвариантными аминокислотами с гидрофобными боковыми заместителями. Гипервариабельные участки Vl, и Ун-областей сближены и образуют доступную растворителю относительно плоскую область связывания антигена.
В Fc-фрагменте lgG Сн2-домены тяжелых цепей ориентированы друг к другу трехцепочечными р-слоями, но непосредственно друг с другом не контактируют из-за ограничений, накладываемых наличием межцепочечных дисульфидных связей в Н2-конце Су2-доменов; между ними находятся углеводные фрагменты, ковалентно связанные с остатками Asn-297 каждой Ц-цепи. Они состоят главным образом из остатков глюкозы, глюкозамина, фруктозы и сиаловой кислоты. Содержание углеводов и состав существенно отличаются у разных классов. Их присоединение является постсинтетическим процессом и происходит упорядоченно во время внутриклеточного транспорта молекулы иммуноглобулина. До сих пор биологическая роль углеводного компонента не ясна.
СуЗ-домены соседних Н-цёпей IgG обращены друг к другу четырехцепочечными р-слоями, и характер их взаимодействия аналогичен связи между Cvl и Cl. Небольшая область контакта между Ст2 и СуЗ-доменами охватывает около 12 остатков.
В молекулах IgM взаимодействие Су2-доменов, по-видимому, происходит по типу Fab—С, т. е. Су2-домены сближены и контактируют четырехцепочечными р-слоями. Наличие дисульфид-ного мостика на С-конце См2-домена способствует тесному сближению N-концов СмЗ-доменов, в результате чего непосредственного контакта между СмЗ-доменами не происходит.
Между Сд1 и Сц2-доменами Н-цепей IgG и IgA и между Сн2 и СнЗ-доменами тяжелых цепей иммуноглобулинов других классов расположен так называемый «шарнирный» участок, создающий высокую степень подвижности между двумя Fab- и Fc-фраг-ментами. Длина «шарнирного» участка иммуноглобулинов разных классов и подклассов варьирует от 15 до 65 а. о. Степень гомологии последовательности «шарнирных» участков составляет 60— 70%. Имеется зависимость между строением «шарнирного» участка и подвижностью Fab-фрагментов подклассов IgG-человека. Наибольшей гибкостью обладает молекула IgG3, у которой «шарнирный» участок, состоящий из,65 а. о., представляет довольно жесткий стержень с гибкими концами, примыкающими к Fab-фрагмен-там. У IgM4, обладающего наиболее жесткой структурой, «шарнирный» участок содержит всего 13 остатков, среди которых 5 остатков Pro и 2 — Cys, образующих межцепочечные дисульфидные связи, IgGl и IgG2 занимают промежуточное положение.
3. Антигенсвязывающие центры антител
Согласно рентгеноструктурным исследованиям комплексов Fab-фрагментов с антигенами связывание антигена происходит в доступной растворителю щели активного центра, образованной вариабельными доменами в N-концевой части легкой и тяжелой цепей. Длина щели антител варьирует от 0,4 до 3,4 нм, а средние размеры области связывания для полимерных антигенов различной природы составляют нм. С антигеном частично контактируют гипервариабельные участки Н- и L-це-пей, расположенные в местах изгибов полипептидной цепи, а также некоторые из аминокислотных остатков, более глубоких внутренних областей цепи. Особую роль в построении анти-генсвязывающего центра антител играет третий гипервариабельный участок Н-цепи, включающий от 1 до 20 а. о. Длина этого участка и контактирующего с ним первого гипервариабельного участка L-цепи во многом определяют размеры активных центров антител.
Способностью связывать антигены с той же эффективностью, что и нативные молекулы антител, обладают Fab- и Р2-фраг-менты иммуноглобулинов. Изолированные Н- и L-цепи имеют весьма низкое сродство к антигену, однако способность к специфическому узнаванию соответствующих антигенов у них сохраняется.
Во взаимодействии с антигеном участвует большое количество аминокислотных остатков обеих цепей молекулы антител, хотя, как правило, большая функциональная нагрузка приходится на Н-цепи.
Основным принципом организации антигенсвязывающих центров иммуноглобулинов является полицентровая структура. Малые антигенные детерминанты связываются на ограниченном участке активного центра, комплементарном данной детерминанте. Большие детерминанты могут занимать практически всю область связывания.
Убедительные доказательства существования пространственно разделенных участков связывания антигена были получены для белка миеломы мыши IgA МОРС 460, конкурентно взаимодействующего с 2,4-динитрофенолом и 2-метил-1,4-нафтохиноном:
--> ЧИТАТЬ ПОЛНОСТЬЮ <--