Контрольная работа: Ветеринарное акушерство
Какова сущность процесса оплодотворения
Оплодотворение — физиологический процесс, заключающийся в слиянии яйца и спермиев с последующей их ассимиляцией и диссимиляцией, в результате чего образуется новая клетка (зигота), обладающая двойной наследственностью.
Во время оплодотворения происходят взаимная ассимиляция и диссимиляция яйца и спермия, в результате которых уже не существует ни яйца, ни спермия, а образуется новая, третья клетка — зигота (от греч. zygotos — соединенный вместе), не тождественная ее двум первоисточникам. Поэтому широко применяемое в биологии называние зиготы оплодотворенным яйцом не соответствует действительности.
Сложный процесс оплодотворения происходит в верхней трети яйцепровода. Спермии быка непосредственно перед оплодотворением должны не менее 6 ч находиться в абдоминальной части яйцепровода. За это время они "созревают" и становятся способными оплодотворять. Такое явление называется капацитацией (инкубацией) спермы. У спермиев барана капацитация длится 1—1,5ч, а у хряка—2—3 ч. Оплодотворение может произойти только при достаточном скоплении спермиев. Количество их значительно колеблется даже у одного и того же вида животных- Спер-
мии ожидают яйцеклетку. Последняя, как и спермии, должна "дотреть". Этот процесс также происходит в яйцепроводе, куда яйцеклетка попадает в стадии овоцита. Там она при наличии активных спермиев выделяет направительные тельца и созревает. Сколько времени требуется для "дозревания" яйцеклетки, пока неясно. У овец, видимо, не более 2ч.
Установлено, что для нормального процесса оплодотворения, а стало быть, получения жизнеспособного плода необходимо соединение только полноценных половых клеток. Неполноценные гаметы не вступают в процесс оплодотворения или вступают, но зародыши гибнут на ранней стадии. В связи с этим важное значение имеют время осеменения, возраст половых клеток и активность спермиев.
Оплодотворение у животных состоит из нескольких стадий.
Первая стадия — денудация—характеризуется тем, что яйцеклетка, проходя по яйцепроводу, освобождается от окружающих ее фолликулярных клеток (лучистого венца). Основная роль в денудации принадлежит спермиям, которые, внедряясь между клетками фолликулярного эпителия, выделяют ферменты, главным образом гиалуронидазу. За счет этих ферментов и неизвестных еще факторов разжижается гиалуроновая кислота, входящая в состав студенистого вещества, связывающего клетки лучистого венца. Рассеивание клеток лучистого венца — это не видовая особенность, и оно может происходить под влиянием спермиев животных другого вида. Так, при осеменении свиней смесью спермы быка и хряка спермии быка освобождают яйцо от фолликулярных клеток, а проникают в него только спермин хряка. Процессу денудации способствуют также механические препятствия, обусловленные ворсинками слизистой оболочки яйцепровода.
Для оплодотворения не обязательно полное освобождение яйцеклетки от клеток лучистого венца. Достаточно лишь места, чтобы спермии проникли через прозрачную оболочку яйцеклетки в околожелточное пространство. Этот процесс, составляющий вторую стадию оплодотворения, в видовом отношении более специфичен. В это время заканчивается созревание яйцеклетки (выделение второго направительного тельца). Ядро, содержащее гаплоидное число хромосом, превращается в женский пронуклеус.
На третьей стадии оплодотворения один, реже несколько спермиев проникают через желточную оболочку яйцеклетки в ее цитоплазму. Это уже строго специфический процесс, поскольку в яйцеклетку могут проникать только спермин своего вида. При этом проникает не весь спермии, а лишь его головка и шейка. Внедрившись в цитоплазму яйцеклетки, спермии претерпевает большие изменения. Головка спермия быстро увеличивается в размере в десятки раз; вследствие ассимиляции цитоплазмы яйца она достигает величины ядра яйцеклетки и превращается в мужской пронуклеус, имеющий, как и женский пронуклеус, половинный набор хромосом.
На четвертой стадии пронуклеусы (ядра яйцеклетки и спермия) постепенно сближаются, вступают в тесный контакт, быстро уменьшаются в объеме и полностью сливаются. Этот процесс, основной в оплодотворении, является еще недостаточно изученным. Образуется качественно новая клетка
(зигота), ядро которой содержит диплоидное число хромосом. Образовавшаяся зигота, таким образом, имеет двойную наследственность. Она получает могучий стимул для дальнейшего развития и начинает быстро дробиться. После первого деления зиготы образуются два бластомера (дочерние клетки); оба бластомера дают четыре внучатых, а четыре внучатых— восемь правнучатых бластомеров и т.д. У млекопитающих такой закономерности нет: количество бластомеров у них даже в начальных стадиях не всегда бывает четным. В отличие от деления одноклеточных организмов клетки, образующиеся в результате деления зиготы, не расходятся, а остаются соединенными; в первый период общая величина зародыша не изменяется, поэтому деление зиготы называется дроблением, Дробление зиготы коровы длится около 8сут. В течение первых 4 сут. дробление происходит в яйцепроводе, а затем в одном из рогов матки. Уже через 48 ч после овуляции зигота состоит из двух бластомеров, через 72 ч — из трех, через 84 ч — из шести и через 96 ч она поступает в стадии морулы в рог матки. Зигота по размеру равна яйцевой клетке или незначительно пре вышает ее, так как с каждым дроблением при увеличении количества бластомеров величина каждого из них соответственно уменьшается.
Наружный слой бластомеров, прилегающий к прозрачной оболочке, называется трофобластом (питающим листком); бластомеры, заключенные в трофобласт, образуют эмбриобласт (зародышевый листок).
Физиология родов у коров
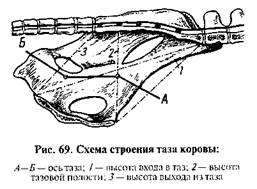
Роды у коровы. Таз коровы по сравнению с тазом животных других видов менее благоприятен для родового акта, Подвздошные кости поднимаются вверх почти под прямым углом, вследствие чего вертикальный диаметр таза располагается под 3—4-м крестцовым позвонком. Вход и таз имеет форму сплюснутого с боков овала. Поперечный диаметр тазовой полости значительно меньше среднего поперечного диаметра входа в таз. Седалищные гребни представляют собой костные пластинки, поднимающиеся кверху и составляющие значительную часть боковых стенок. Лишь сравнительно небольшой участок последних образован связками таза. Выход из таза с боков ограничен седалищными буграми в виде костных пластин, зажимающих плод при его продвижении кзади. Наконец, вогнутая поверхность крестца и неровное, с углублением, дно тазовой полости придают оси таза форму ломаной линии (рис, 69). Плод имеет максимальную ширину головы в области лобных костей; у него более короткие конечности, чем у жеребенка- Стадия раскрытия шейки матки продолжается до 12 ч. Корова ведет себя более спокойно, чем кобыла. Изредка продолжительно тихо мычит. Под влиянием сокращения матки через раскрытую шейку выпячивается плодный пузырь, состоящий из сосудистой и околоплодной оболочек или алланто-хориона. Плодный пузырь может выпячиваться за пределы вульвы и затем разрываться, но чаше плодные оболочки разрываются еще во влагалище и из родовых путей выступают предлежащие конечности.
Стадия выведения плода длится от 20 мин до 3—4 ч и больше. Такая большая продолжительность обусловлена комплексом причин. Вследствие не косого, а почти прямого положения входа в таз плод сразу всей массой предлежащих органов приходит в соприкосновение с его верхней, нижней и боковыми стенками; далее, попав в полость таза, предлежащие органы располагаются под вогнутостью тела крестцовой кости и углублении дна таза. В этом же месте плод сбоку зажимается мощными седалищными гребнями. При продвижении к выходу плод встречает тройное сопротивление, а именно: со стороны опускающегося в просвет таза каудального конца крестцовой кости, возвышающимся перед ним дном тазовой полости, и, наконец, боковых стенок выхода таза, образованных костными пластинками седалищных бугров. Последнее препятствие обычно и служит наиболее частой причиной задержки родового процесса уже в тот момент, когда плод вошел в тазовую полость, а из вульвы выступают конечности и лицевая часть черепа. В этих случаях неопытные практики, оказывай помощь, нередко не учитывают особенностей структуры таза. Укрепив петли за конечности и голову, они начинают вытягивать плод по направлению оси туловища. Такой прием в большинстве случаев безрезультатен, так как плод встречает непреодолимое препятствие в виде седалищных бугров. В то же время достаточно небольшого натяжения наверх и назад, чтобы предлежащая часть переместилась из участка, ущемленного между костями выхода, в пространство между двумя связками, и роды легко и быстро закончились.
При двойнях второй плод родится спустя некоторое время после первого. В стадии выведения плода корова обычно лежит на животе или на боку. У нее отмечается сильное общее возбуждение.
Последовая стадия у жвачных значительное длиннее, чем у лошади. Нормальным сроком можно считать до 10 ч с момента выведения плода (у большинства коров 5—6 ч). Некоторые практики расценивают как нормальное явление задержание последа у коров до 2—3 и даже до 5 сут. Такое мнение не имеет никаких анатомо-физиологических обоснований. Длительный срок отделения последа у коровы объясняется структурой плаценты. Материнская часть плаценты представлена сочной губчатой тканью, сидящей на ножке и вдающейся в полость матки. Сокращение и ретракция мышц матки только сближают карункулы, а не отжимают плодные оболочки, так как хорион собирается в складки между карункулами. Только после понижения тургора начинается освобождение ворсин хориона из крипт карункула.
Через 2—Зч после родов полость матки оказывается спавшейся. Карункулы заполняют почти весь ее просвет. Матка становится плотной, на ее поверхности прощупываются складки. Рог, служивший плодовместилищем, больше свободного. На 3—5-й день просвет канала шейки сужается настолько, что пропускает только 2—3 пальца; закрытие его заканчивается после прекращения выделения лохи и. Вибрация средних маточных артерий у коров-первотелок после рождения плода продолжается 20 ч- Лохии у коров в первые дни кровянистые; на 3—5-й день они образуют в шейке матки пробку, состоящую из густой, мутной, тягучей слизи. Затем лохии становятся шоколадного цвета; с 8-го дня приобретают характер слизи, а к 10—14-му дню прекращаются. Выделение лохии после 14-го дня — признак заболевания матки.
Инволюция желтого тела беременности заканчивается к 16-му дню послеродового периода и сопровождается интенсивным ростом фолликулов. В процессе рассасывания желтого тела его клетки подвергаются вакуолизации. Их протоплазма содержит эозинофильную зернистость. Вокруг клеток желтого тела замечают лейкоцитарную инфильтрацию. Постепенно количество клеток желтого тела начинает уменьшаться. Сосуды, обильно пронизывающие желтое тело, с уменьшением его общей величины сближаются, образуя густую сеть. Постепенно и сосуды подвергаются дегенерации, превращаясь в соединительнотканные тяжи — основу белого тела.
После отделения последа поверхность карункула покрывается кровяными сгустками; образующие его основу сосуды подвергаются тромбозу. Весь комплекс тканей, принимавших участие в формировании карункула, распадается, превращаясь в однородную массу. В соединительнотканных прослойках скапливается большое количество лейкоцитов.
К 10-му дню вся масса карункула расплавляется, на его месте заметен небольшой сосудистый клубок. В этот период остатки карункула при дотрагивании до них рукой легко отпадают без кровотечения. Следовательно, инволюция карункула сводится к перерождению и распаду его клеток с частичным рассасыванием и отторжением. Там, где был карункул, появляется свободная от эпителия поверхность слизистой оболочки, которая эпителизируется путем нарастания клеточных элементов маточных желез. Не только площадь бывшего карункула, но и участки слизистой оболочки матки, располагающиеся между карункулами, лишаются покровного эпителия и только к 10— 14-му дню эпителизируются тем же путем, что и поверхность карункула.
Продолжительность родового акта зависит от физиологического состояния животных, У коров, пользующихся активным моционом, все стадии родов протекают интенсивнее и быстрее. Важное значение имеет и место проведения самих родов. При родах в стойлах у коров и новорожденных телят, получающих молозиво без учета их физиологической потребности (времени, кратности и количества), нередко возникают различные заболевания. Поэтому целесообразно проводить роды в специальных боксах и содержать телят вместе с матерями первые 4—5 дней. При этом коров нужно обязательно поддаивать. Роды в боксах благоприятно сказываются на течении всех стадий родового акта, состоянии здоровья коров и телят.
Корову в боксе содержат без привязи, она принимает удобное для нее положение при родах, которые протекают в тихой и спокойной обстановке. Корова многократно и тщательно облизывает всего теленка (необходимость обязательного облизывания коровой новорожденного доказал П. А. Флегматов, выявив мистические свойства околоплодной жидкости), что усиливает деятельность сердечнососудистой, дыхательной, пищеварительной и других систем.
Облизывание в сочетании с актом сосания теленка стимулирует сокращение матки, в результате чего ускоряется отделение последа и сокращается время инволюции матки. Телята, родившиеся в боксе, встают раньше. У них быстрее и вовремя путем подсоса реализуется пищевой рефлекс, что имеет важное физиологическое значение в повышении иммунной резистентности организма новорожденного. У таких телят раньше отходит и первородный кал. Каждый раз после перевода родивших коров в индивидуальные стойла послеродовой секции боксы тщательно очищают, моют и дезинфицируют.
При отсутствии подсоса корову первый раз доят через 30—40 мин после рождения теленка. Это полезно и для матери, и для новорожденного. Чем раньше теленок (жеребенок, ягненок, поросенок) получит молозиво, тем меньше у него случаев заболеваний желудочно-кишечного тракта. Новорожденному выпаивают 1,5—2л молозива, лучше всего через специальные соски. Оставшееся молозиво выпаивают корове что способствует усилению моторной функции матки и более быстрому отделению последа.
Всех коров независимо от продуктивности в первые 7—10 дней надо доить не реже 4—5 раз в сутки, чтобы вовлечь в лактацию все доли вымени.
В первые 5—7 дней коров следует кормить только хорошим сеном, а затем постепенно включать в рацион сочные корма и концентраты. Преждевременный перевод животных на полный рацион (раньше чем через 2 нед.) задерживает инволюцию матки.