Курсовая работа: Биосинтез мембранных белков и их встраивание в биомембрану
Для удаления временных N-концевых сигнальных пептидов необходимы специфические белки. Наиболее полно охарактеризованы сигнальные протеазы из Е.coli. Большинство экспортируемых белков Е.coli содержат сигнальный пептид, который отщепляется на периплазматической поверхности внутренней мембраны с помощью лидер-пептидазы; ее структура представлена на рис.2.
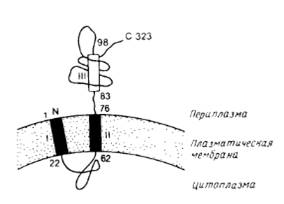
Рис.2. Предполагаемая топология лидер-пептидазы из Е.coli.
Аминокислотные остатки пронумерованы. Прямоугольниками обозначены гидрофобные сегменты. Первый гидрофобный участок не был обнаружен экспериментально, вывод о его существовании был сделан исходя из его гидрофобности. Наличие и ориентация второго гидрофобного участка установлены экспериментально и, по-видимому, он функционирует как внутренняя сигнальная последовательность. Третий гидрофобный сегмент имеет слабо выраженный гидрофобный характер и является частью периплазматического домена.
Для переноса белков через внутреннюю мембрану эта пептидаза не нужна, но она необходима для высвобождения экспортируемого белка из цитоплазматической мембраны. Invitro очищенный фермент мог функционировать, будучи включенным в липосомы. Специфичность расщепления весьма высока, но не определяется исключительно аминокислотной последовательностью вблизи сайта расщепления. Сигнальная пептидаза, функционирующая в эндоплазматическом ретикулуме, имеет ту же специфичность, что и соответствующий фермент Е.coli, что неудивительно, если учесть сходство сигнальных последовательностей. Была очищена сигнальная пептидаза из микросом эукариот. Показано, что она ассоциирована с другими полипептидами, возможно имеющими отношение к механизму переноса.
У Е.coli имеется вторая сигнальная пептидаза, участвующая в процессинге пролипопротеинов. Эти полипептидные компоненты оболочки Е.coli замечательны тем, что при созревании их N-конец модифицируется с помощью глицерида. Пролипопротеиновая сигнальная пептидаза также находится в цитоплазматической мембране. После отщепления сигнальный пептид остается в цитоплазматической мембране и разрушается с помощью мембраносвязанного фермента протеазы ΙV.
В митохондриях и хлоропластах должно присутствовать несколько сигнальных пептидаз, поскольку процессинг происходит более чем в одном компартменте. Растворимую пептидазу из митохондриального матрикса удалось частично очистить, но охарактеризована она не полностью.
4. Растворимые и мембраносвязанные белки, необходимые для
переноса
Идентифицировано несколько цитозольных и мембраносвязанных белковых компонентов, необходимых для переноса. Наиболее детально охарактеризованы белковые факторы, участвующие во встраивании белков в эндоплазматический ретикулум млекопитающих (рис.3).
1. Сигнал-распознающая частица (СРЧ). Это растворимый рибонуклеопротеиновый комплекс, состоящий из шести разных белков и молекулы 7S-РНК. СРЧ необходима для инициации переноса. Она связывается с сигнальной последовательностью образующегося полипептида во время его синтеза на рибосоме. Для препролактина, например, константа диссоциации составляет 1нМ. С помощью метода фотохимического сшивания был идентифицирован один из полипептидов (54 кДа), непосредственно взаимодействующий с сигнальной последовательностью предшественника. По некоторым данным, полученным для бесклеточных систем, связывание СРЧ ингибирует трансляцию или вызывает ее задержку. Впрочем, не исключено, что этот феномен является артефактом; во всяком случае, как было показано на модельных опытах, его не обязательно привлекать для объяснения кинетики переноса белков invivo. Одна из вероятных функции СРЧ состоит в предотвращении неправильного свертывания образующегося полипептида, которое может блокировать перенос (например, из-за экранирования сигнальных последовательностей). Задержка трансляции должна уменьшать вероятность такого ошибочного свертывания и, следовательно, увеличивать эффективность переноса белков.
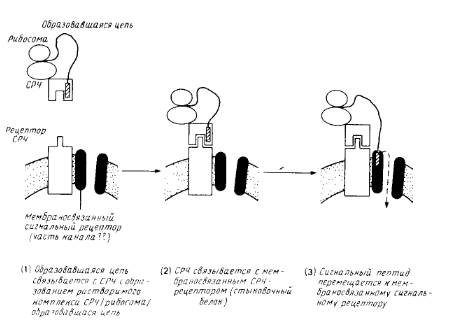
Рис.3. Схематическое изображение ранних стадий котрансляционного переноса полипептида через эндоплазматический ретикулум млекопитающих.
Сигнал-распознающая частица и СРЧ-рецептор (стыковой белок) хорошо охарактеризованы. Мембраносвязывающий сигнальный рецептор изображен как компонент канала через мембрану, но существование таких каналов и роль сигнального рецептора не бесспорны. После образования комплекса между сигнальным пептидом и мембраносвязанным рецептором на стадии 3 СРЧ и стыковочный белок могут диссоциировать и принимать участие в новом цикле, оставляя мембраносвязанную рибосому и образовавшуюся цепь присоединенными к аппарату переноса.
Некоторые небольшие белки (<8,5 кДа) транспортируются в эндоплазматический ретикулум независимо от СРЧ. В их число входят препропептид GLaлягушки, препромеллитин (оба они являются предшественниками секретируемых белков) и пробелок оболочки фага М13. Во всех этих примерах конформация предшественника такова, что белки должны оставаться способными к переносу даже в отсутствии СРЧ и рибосом.
2. Рецептор СРЧ, или стыковочный белок. Комплекс СРЧ/рибосома/образующаяся полипептидная цепь транспортируется в шероховатый эндоплазматический ретикулум, преодолевая энергию сильного взаимодействия между СРЧ, называемым также стыковочным белком. Рецептор СРЧ содержит субъединицу с молекулярной массой 73 кДа, присоединенную N-концом к мембране. Вероятно, рибосома также связывается со специфическими рецепторами, присутствующими в мембране.
3. Рецептор сигнальной последовательности. Сигнальная последовательность на образующейся полипептидной цепи перемещается от СРЧ ко второму рецептору, находящемуся в мембране и называемому рецептором сигнальной последовательности. Об этом свидетельствуют результаты опытов по фотохимическому сшиванию, в которых используется метка, связанная с сигнальной последовательностью препролактина. Предполагаемый мембраносвязанный рецептор представляет собой гликопротеин с молекулярной массой 35 кДа. Возможно, он образует часть канала, через который осуществляется перенос. С помощью такого же подхода с использованием поперечной сшивки и синтетического сигнального пептида был обнаружен еще один кандидат на роль рецептора сигнальной последовательности (45 кДа). Связь между этими двумя белками неизвестна и функции их до конца не установлены. Как только образовавшаяся полипептидная цепь связывается с мембраносвязанным рецепторм, СРЧ и ее рецептор могут освободиться от рибосомы и принять участие в новом цикле. О предполагаемом канале, участвующем в переносе, ничего неизвестно; очистка его является довольно сложной задачей.
5. Сборка мультисубъединичных комплексов и обновление
мембранных белков
После встраивания мембранного полипептида в мембрану он еще должен приобрести правильную конформацию, обеспечивающую его биологическую активность, а если речь идет о мультисубъединичных комплексах, то связаться с другими белками. В частности, у эукариот при этом должны произойти различные ковалентные модификации, например гликозилирование, ацилирование, сульфирование или образование дисульфидных связей. Даже когда такие модификации не являются необходимыми, процесс конформационного созревания может быть медленным и отстоять по времени от встраивания в мембрану.
Например, у Е.coli четко наблюдается сборка стабильных тримеров обоих белков, LamB и OmpF, после включения соответствующих мономеров в наружную мембрану, при этом созревание LamB занимает около 5 мин. В эукариотических клетках гликопротеин гемагглютинина вируса гриппа, прежде чем попасть из эндоплазматического ретикулума в комплекс Гольджи, должен сформировать правильную четвертичную структуру, соответствующую зрелой форме. Несвернутые молекулы гемагглютинина остаются в эндоплазматическом ретикулуме. Образование тримеров занимает примерно 7-10 мин. Сходная олигомеризация наблюдается также для G-белка вируса везикулярного стоматита.
Сборка многих субъединичных комплексов, содержащих разные субъединицы, тоже, по-видимому, происходит в эндоплазматическом ретикулуме. Примером служит никотиновый ацетилхолиновый рецептор, который содержит две α-субъединицы и по одной β-, γ- и δ-субъединице (рис.4).
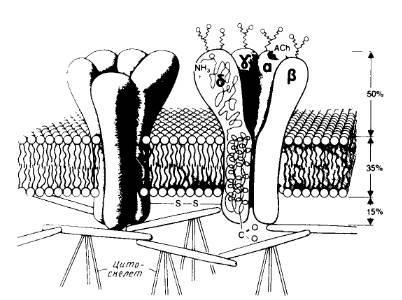
Рис.4. Модель канала никотинового ацетилхолинового рецептора.
Показаны общий вид канала и его расположение в мембране, отмечены участки гликозилирования с наружной стороны и места связывания ацетилхолина на α-субъединицах.
С помощью антител можно различить отдельные формы α-субъединицы: 1) начальный продукт, встраивающийся в эндоплазматический ретикулум; эта форма не может связывать антагонист α-бунгаротоксин; 2) форма, способная связываться с α-бунгаротоксином и образующаяся через несколько минут после завершения трансляции в эндоплазматическом ретикулуме; 3) зрелый рецептор, содержащий все субъединицы (α2 βγδ), который обнаруживается через 15мин после завершения трансляции в эндоплазматическом ретикулуме; 4) готовый рецептор на клеточной поверхности, появляющийся спустя примерно 2ч после трансляции. Созревание включает образование дисульфидных связей, олигосахаридный процессинг и ацилирование при участии жирных кислот. Вероятно, определенную роль в сборке, происходящей в комплексе Гольджи, играет фосфорилирование субъединиц.
Решающим фактором процесса сборки является, вероятно, стабильность таких стехиометрических комплексов, как ацетилхолиновый рецептор. По-видимому, в некоторых системах отдельные субъединицы синтезируются в значительном избытке и не образуют стабильных комплексов, а подвергаются протеолитическому расщеплению.
Изучался также процесс созревания и сборки структуры, образующей Na-канал. Необходимым условием созревания является образование дисульфидной связи между α- и β2 -субъединицами, однако, это событие происходит спустя примерно 1ч после трансляции и транспорта субъединиц в аппарат Гольджи, а рецептор появляется на клеточной поверхности через 4ч после трансляции. В этом случае свободные α- субъединицы не подвергаются быстрой деградации, а сохраняются в межклеточном пуле и, возможно, используются в дальнейшем в качестве предшественников для формирования канала в растущих нейронах.
Созревшие мембранные белки подвергаются непрерывному обновлению. Период полуобновления Na-канала составляет около 30ч, что типично для поверхностных белков. Обновление большой субъединицы Na/К-АТРазы в растущих клетках в культуре происходит за время 20-40ч. По-видимому, деградация по крайней мере некоторых белков происходит в лизосомах.