Курсовая работа: Гипотеза мира РНК
После удлинения выступающей цепи ДНК до конца матрицы происходит транслокация, т.е. перемещение матрицы и белковых субъединиц фермента на заново синтезированный конец теломеразной ДНК, и весь цикл повторяется вновь. После завершения удлинения одноцепочечной З'-концевой теломерной последовательности вторая цепь ДНК (С-цепь) достраивается с помощью обычной ДНК-полимеразы. Таким образом происходит решение проблемы концевой репликации ДНК у эукариот.
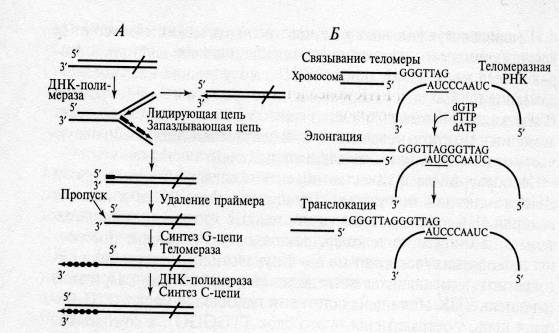
Рис. 1. Репликация теломерных участков эукариотических хромосом (Цитировано по [1]):
А — возникновение недореплицирования 5'-конца линейной хромосомы и синтез на этом концевом участке теломерной ДНК с помощью теломеразы;
Б — основные этапы синтеза теломерного повтора теломеразой
I.1.2 Механизм обратной транскрипции
На обратной транскрипции основано размножение ретровирусов (вирусы, у которых геномом служит не ДНК, как обычно, а РНК) и ретротранспозонов (являются транспозиционными элементами, которые не имеют вирионных частиц, и, следовательно, в отличие от ретровирусов, не могут независимо «переносить себя» между клетками), образование так называемых ретропсевдогенов (или процессированные псевдогены это ретропоследовательности, которые потеряли свою функцию, они несут все признаки функциональных ретропоследовательностей, но имеют молекулярные дефекты, которые не дают им экспрессироваться)и достройка кончиков хромосом (теломер), укорачивающихся при каждом клеточном делении.Если молекула ДНК повреждена — например, подверглась разрыву (double-strand break, DSB) — для ее починки необходима матрица, в которой последовательность нуклеотидов соответствует исходному, «правильному» состоянию поврежденного участка. Ранее считалось, что в качестве таких матриц всегда используются другие молекулы ДНК. Позже было установлено, что иногда эти ДНК-матрицы синтезируются путем обратной транскрипции на основе РНК при участии ретротранспозонов.
При изучении ретровирусов, геном которых представлен молекулами одноцепочечной РНК, было обнаружено, что в процессе внутриклеточного развития ретровирус проходит стадию интеграции своего генома в виде двухцепочечной ДНК в хромосомы клетки-хозяина. В 1964 г. Темин выдвинул гипотезу о существовании вирусспецифичного фермента, способного синтезировать на РНК-матрице комплементарную ДНК. Усилия, направленные на выделение такого фермента, увенчались успехом, и в 1970 г. Темин с Мизутани, а также независимо от них Балтимор открыли искомый фермент в препарате внеклеточных вирионов вируса саркомы Рауса. Данная РНК-зависимая ДНК-полимераза получила название обратная транскриптаза, или ревертаза.
Каждый вирион(полноценная вирусная частица, состоящая из нуклеиновой кислоты и белковой оболочки) ретровирусов содержит две идентичные цепи РНК размером от 8000 до 10 000 нуклеотидов. Области 5'- и 3'-концов обеих цепей модифицированы, как у всех эукариотических мРНК (5'-кэпы, З'-полиадениловые хвосты). Вирусные РНК имеют 5 структурных элементов: 1) прямые повторы на 5'- и З'-концах РНК (R); 2) последовательность из 80 — 120 нуклеотидов, находящуюся около 5 концевого повтора (U5); 3) последовательность из 170 — 1200 нуклеотидов около З'-концевого повтора (U3); 4) последовательность из 15 20 нуклеотидов (Р), в пределах которой клеточная тРНК комплементарно взаимодействует с ретровирусной РНК, что создает праймер для синтеза первой цепи ДНК; 5) сегмент Pu, находящийся непосредственно перед повтором U3 и являющийся сайтом для праймирования второй цепи ДНК — такой сегмент одинаков у РНК всех ретровирусов определенного типа.
Этапы обратной транскрипции:
1. Наращивание тРНК-праймера на матрицах U5 и R в направлении 3'→5'. Роль РНК-праймера выполняет одна из клеточных тРНК (например, триптофановая, пролиновая и т.д.). На расстоянии примерно 100 — 200 нуклеотидов от 5'-конца РНК (для каждого вируса — это величина постоянная) имеется участок, комплементарный З'-концевой последовательности молекулы тРНК, который используется в качестве затравки. Этот участок обычно обозначают как pbs (от англ.primerbindingsite участок связывания затравки). Обратная транскриптаза синтезирует сегмент ДНК, комплементарный 5'-концевой последовательности вирусной РНК. Этот сегмент принято называть (-) «strong-stop» ДНК, поскольку синтез ДНК после завершения копирования 5'-конца матрицы временно останавливается. (-) «strong-stop» ДНК содержит последовательности, комплементарные концевому району R и району U5. Таким образом, синтез ДНК начинается недалеко от 5'-конца матрицы и образуется короткий продукт. Но этот короткий продукт (-) «strong-stop» имеет последовательность, комплементарную также и З'-концу вирусной РНК, а как известно, для снятия ДНК-копии с З'-конца матрицы всегда требуется праймер. У ретровирусов комплемент З'-конца матрицы производится в «удобном» месте, а затем переносится на «свое» место. Это происходит следующим образом: 5'-конец вирусной РНК, образующий дуплекс с (-) «strong-stop» ДНК, разрушается под влиянием присущей обратной транскриптазе активности РНКазы Н.
2. РНКаза Н, специфичная к РНК в составе гибридного РНК-ДНК дуплекса, расщепляет сегмент РНК этого дуплекса. В результате (-) «strong-stop» (RU5) оказывается в однонитевой форме и может взаимодействовать с З'-концом (с участком R) той же самой или другой молекулы вирусной РНК, поскольку на З'-конце РНК имеется повтор R..
3. Новосинтезированная короткая цепь ДНК вместе с праймером «перепрыгивает» на З'-конец матрицы и взаимодействует там с комплементарным ей участком К.
4. Цепь ДНК удлиняется, в качестве матрицы используется остальная часть вирусной РНК. На этой стадии в качестве затравки выступает уже (-)«strong-stop» ДНК; элонгация затравки приводит к синтезу (-) цепи ДНК, в которой отсутствует комплемент района RU5, поскольку соответствующий участок (+) матрицы был разрушен РНКазой Н.
5. К моменту завершения синтеза первой цепи ДНК большая часть вирусной РНК разрушается РНКазой Н.
6. Синтез З'-конца второй цепи ДНК.
7. Удаление тРНК и оставшегося участка вирусной (+) РНК РНКазой Н.
8. Второй прыжок, в результате которого новосинтезированная вторая цепь ДНК комплементарно взаимодействует с тРНК-связывающей последовательностью первой цепи.
9. Удлинение З'-концов каждой цепи, образование дуплекса ДНК.
Вся последовательность реакций протекает без явного участия ферментов репликации клетки-хозяина (топоизомеразы, хеликазы, праймазы, ДНК-связывающего белка, лигазы и т.д.). При этом следует отметить, что молекулы вирусных ДНК длиннее молекул вирусных РНК, которые послужили матрицей для обратной транскрипции. Действительно, к 5'-концу (+) цепи вирусной ДНК добавилась последовательность U3, а к 3 -концу этой цепи — последовательность U5. В результате на концах молекулы вирус специфической ДНК появился длинный (несколько сотен нуклеотидов) концевой повтор (ДКП или LTR.), имеющий структуруU3U5.
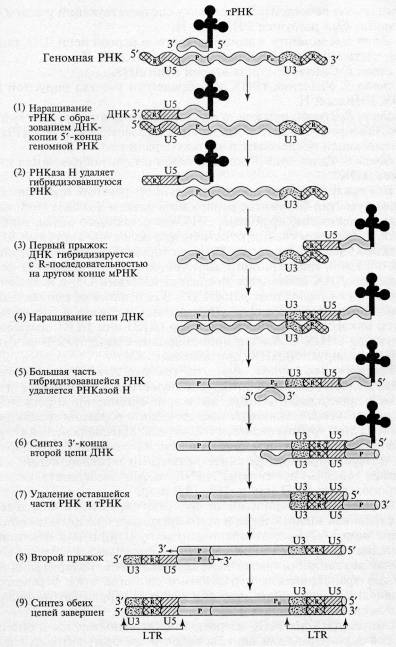
Рис.2. Схема обратной транскрипции ретровирусной РНК с образованием двуцепочечной ДНК (DarnellJ., et.al. MolecularCellBiology. – N. Y.: ScientificAmer. Books, 1986. – P. 1052)
Синтез ДНК на РНК-матрице invitroи ревертаза используется в генетической инженерии для синтеза генов и их фрагментов, а также целенаправленного синтеза на матричных РНК комплементарных молекул ДНК (кДНК) для расшифровки первичной структуры РНК и белков.
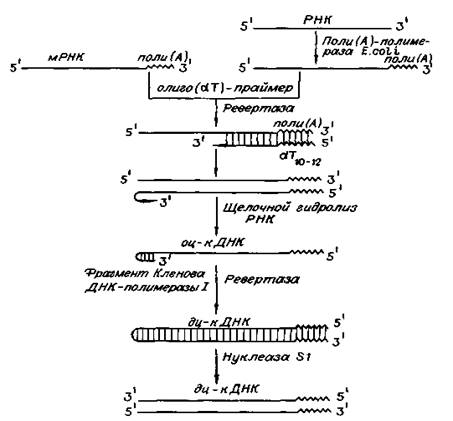
Рис. 3. Схема получения кДНК с использованием ревертазы вируса и трех дополнительных ферментов: поли (А)-полимеразы, фрагмента Кленова ДНК-полимеразы I и нуклеазы S1. (Цитировано по)
Реакцию обратной транскрипции проводят в специально подобранных условиях с использованием сильных ингибиторов РНКазной активности. При этом удается получать полноразмерные ДНК-копии целевых молекул РНК. В качестве праймера при обратной транскрипции поли (А) - содержащих мРНК используют олигo (dT)-праймер, а для молекул РНК, не имеющих З'-поли (А) концов, — химически синтезированные олигонуклеотиды, комплементарные З'-концу изучаемой РНК. После синтеза на мРНК комплементарной цепи ДНК и разрушения РНК (обычно применяют обработку щелочью) осуществляют синтез второй цепи ДНК. При этом используют способность ревертазы образовывать на 3'-концах одноцепочечных кДНК самокомплементарные шпильки, которые могут выполнять функции праймера. Матрицей служит первая цепь кДНК. Данная реакция может катализироваться как ревертазой, так и ДНК-полимеразой I E. coli. Показано, что сочетание этих двух ферментов позволяет повысить выход полноценных двухцепочечных молекул кДНК. По окончании синтеза первая и вторая цепи кДНК остаются ковалентно связанными петлей шпильки, служившей праймером при синтезе второй цепи. Эту петлю расщепляют эндонуклеазой S1, специфически разрушающей одноцепочечные участки нуклеиновых кислот. Образующиеся при этом концы не всегда оказываются тупыми, и для повышения эффективности последующего клонирования их репарируют до тупых с помощью фрагмента Кленова ДНК-полимеразы I E. сoli (остающаяся часть молекулы, которая сохраняет присущие ей каталитические активности).
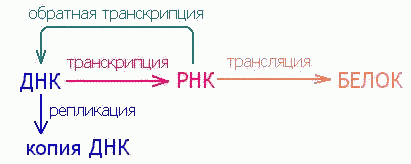
Уже одно это открытие формально поставило РНК в центр основного постулата молекулярной генетики, так как показало, что поток генетической информации распространяется от РНК не в одном, а в двух направлениях: не только к белку, но и к ДНК. Все более глубокое проникновение в механизмы основных молекулярно-генетических процессов (репликацию, транскрипцию и трансляцию) способствовало возникновению понятия о неканонических функциях РНК, осознанию полифункциональности рибонуклеиновых кислот.
I.2 Функциональные возможности РНК