Курсовая работа: Креатинфосфокиназа
Можно показать, что член keравен максимальной скорости. При выводе уравнения (11) не было сделано допущения о влиянии, которое может оказывать взаимодействие фермента с одним из субстратов на взаимодействие с другим.
Если каждый субстрат взаимодействует только со своим участком связывания и не оказывает влияния на сродство фермента к другому субстрату, то KS A = KM A и KS B = KM B ; уравнение (11) при этом упрощается до
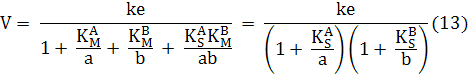
Таким образом, обе константы Михаэлиса можно определить просто путем изменения концентрации одного из субстратов при постоянной концентрации другого. ( Диксон М., Уэбб Э., Ферменты, Т. 1, М., 1982, с. 134, 135).
Перенос энергии из митохондрий в цитоплазму клетки миокарда с помощью изоферментов КФК
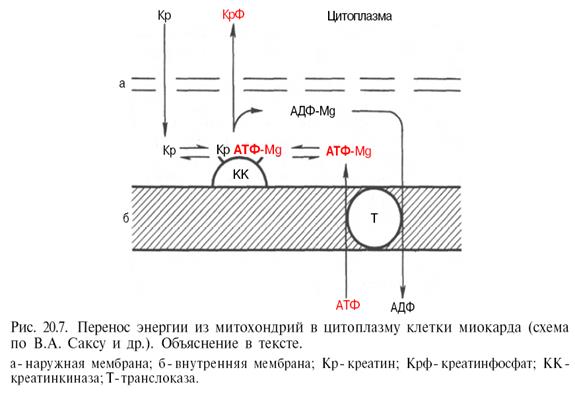
В последнее время появились данные, доказывающие, что креатинфосфат в мышечной ткани (в частности, в сердечной мышце) способен выполнять не только роль депо легкомобилизуемых макроэргических фосфатных групп, но также роль транспортной формы макроэргических фосфатных связей, образующихся в процессе тканевого дыхания и связанного с ним окислительного фосфорилирования. Предложена схема переноса энергии из митохондрий в цитоплазму клетки миокарда (рис. 20.7).
АТФ, синтезированный в матриксе митохондрий, переносится через внутреннюю мембрану с участием специфической АТФ-АДФ-транслоказы на активный центр митохондриального изофермента креатинкиназы, который расположен на внешней стороне внутренней мембраны; в межмембранном пространстве (в присутствии ионов Mg2+ ) при наличии в среде креатина образуется равновесный тройной фермент-субстратный комплекс креатин–креатинкиназа–АТФ-Mg2+ , который затем распадается на креатинфосфат и АДФ Mg2+ . Креатинфосфат диффундирует в цитоплазму, где используется в миофибриллярной креатинкиназной реакции для рефосфорилирования АДФ, образовавшегося при сокращении. Высказываются предположения, что не только в сердечной мышце, но и в скелетной мускулатуре имеется подобный путь транспорта энергии из митохондрий в миофибриллы. (Березов Т. Т., Коровкин Б. Ф., Биологическая химия, М., 1998, с. 580, 654, 655). Изоферменты КФК обнаружены во всех субклеточных структурах, продуцирующих либо использующих энергию, - в митохондриальной мембране, миофибриллах, мембране саркоплазматического ретикулума (СПР), плазматической клеточной мембране и др. (Фролькис Р. А., Воронков Г. С., Дубур Г. Я., Циомик В. А., Гомон И. В. – Некоторые аспекты изучения активности креатинфосфокиназы в мембранах саркоплазматического ретикулума миокарда в условиях экспериментальной коронарной недостаточности //Бюллетень экспериментальной биологии и медицины, М., 1983, №5, с. 53).
Активность КФК в мембранах саркоплазматического ретикулума миокарда в условиях экспериментальной коронарной недостаточности
Изоферменты КФК находятся в тесном функциональном сопряжении с другими ферментами, обеспечивающими перенос веществ и ионов через мембраны. Так, в митохондриях КФК функционирует в тесном сопряжении с АТФ-АДФ-транслоказой, в миофибриллах, мембране СПР и сарколемме – соответственно с Mg-зависимой, Са-зависимой и Na, К-зависимой АТФ-азой.
Важность изучения роли мембранных образований в процессах транспорта и утилизации энергии клетками миокарда приобретает особое значение в условиях его ишемии, при которой наблюдаются существенные изменения в структуре и свойствах белково-липидного слоя мембран и прежде всего в их фосфолипидном составе. Последнее обстоятельство особенно важно в связи с имеющимися в литературе сведениями о том, что некоторые изоферменты КФК, скорее всего, связываются с мембраной с помощью именно фосфолипидов.
В лаборатории биохимии Украинского НИИ кардиологии им. Н. Д. Стражеско (Киев) и в лаборатории мембраноактивных соединений b-дикетонов Института органического синтеза АН Латвийской ССР (Рига) изучали активность митохондриального изофермента КФК при дозированном ограничении коронарного кровотока , а также вопрос об энергообеспечении транспорта Са 2+ через мембраны СПР клеток миокарда и связи этого процесса с некоторыми другими показателями, определяющими функциональное состояние мембранного аппарата СПР.
Опыты проводили на кроликах в условиях острого ограничения коронарного кровотока (внутривенное введение вазопрессина в дозе 0,5 ед/кг), а также на фоне предварительного введения (за 1 ч до вазопрессина) животным антиоксидантного препарата AV-156Na. В мембранах СПР определяли активность КФК, некоторые показатели перекисного окисления липидов, а также содержание фосфолипидов.
Результаты проведенного исследования (таблица №2) свидетельствуют о выраженном влиянии экспериментальной ишемии на все изучаемые звенья метаболизма миокарда. Так, активность изофермента КФК, связанного с мембраной СПР, снижается при вазопрессиновой коронарной недостаточности более чем на 25%.
Таблица №2. Активность КФК, содержание фосфолипидов в мебранах СПР миокарда кроликов при экспериментальной ишемии
| Условия опытов |
КФК, мкмоль креатинина на 1мг белка за 1ч |
Фосфолипиды, мкмоль Р на 1 мг белка |
| Контроль | 11,90 ± 0,36 | 0,37 ± 0,01 |
|
Вазопрессин Вазопрессин на фоне антиоксиданта |
8,22 ± 0,10 10,28 ± 0,70 |
0,22 ± 0,01 0,42 ± 0,01 |
Сдвиги, обнаруженные со стороны активности КФК, тесно сопряжены со структурными изменениями в мембранах СПР, прежде всего их липидного состава. Так, в условиях коронарной недостаточности наблюдается некоторое снижение содержания мембраносвязанного холестерина, а также резкое падение уровня фосфолипидов (на 40%),
При введении антиоксидантного препарата AV-156Na наблюдалось однонаправленное изменение изучаемых показателей в сторону их нормализации. Этот эффект свидетельствует о том, что уровень протекания перекисных реакций в миокарде во многом определяет характер изменения отдельных метаболических процессов в мембранах.
Обнаруженное снижении активности КФК мембран СПР в условиях экспериментальной ишемии может свидетельствовать об определенном нарушении нормального снабжения энергией транспортной Са-АТФ-азы мембран СПР. При этом изменение активности КФК происходит на фоне значительного снижения в мембранах СПР сердца кроликов уровня фосфолипидов. Возможно, изофермент КФК. Связанный с мембраной СПР, является липидзависимым ферментом и для проявления полной его активности сохранение нормальной липидной структуры мембран – важное и необходимое условие. (Фролькис Р. А., Воронков Г. С. , Дубур Г. Я., Циомик В. А., Гомон И. В. – Некоторые аспекты изучения активности креатинфосфокиназы в мембранах саркоплазматического ретикулума миокарда в условиях экспериментальной коронарной недостаточности //Бюллетень экспериментальной биологии и медицины, М., 1983, №5, с. 53, 54).
Влияние алкогольной интоксикации и процессов старения на активность креатинфосфокиназы и ее изоферментов
В отделе фармакологии Института экспериментальной медицины АМН СССР (Ленинград) проводили опыты по выявлению возрастных изменений активности КФК в сыворотке крови интактных животных в возрасте 4 и 24 месяцев, а также изучали влияние острой алкогольной интоксикации на активность КФК и ее изоферментов в тканях и биологических жидкостях крыс 2 групп: 1) животные в возрасте 4 месяцев, которым за 3 часа до начала эксперимента внутрибрюшинно вводили 25% раствор этанола в дозе 3 и 5 г/кг; 2) контроль – крысы, которым вводили соответствующие объемы физиологического раствора. Перед декапитацией у этих животных под легким эфирным наркозом брали ликвор. В экстракте головного мозга, полученном из коры больших полушарий, ликворе и сыворотке крови определяли активность КФК и ее изоферментный профиль. Общую активность фермента определяли колориметрическим методом. Изоферменты разделяли с помощью хроматографии на колонке на сефадексе ДЕАЕ А50. Количество белка определяли микробиуретовым методом. Статическую обработку проводили по Стьюденту.
Анализируя данные об общей активности КФК в сыворотке крови крыс с острой алкогольной интоксикацией, а также у интактных крыс разного возраста, можно сделать вывод о том, что интактные животные в возрасте 4-5 месяцев по активности КФК и ее изоферментному спектру резко отличаются от интактных крыс в возрасте 24 месяцев (таблица №3).