Курсовая работа: Липиды центральной нервной системы и структура клеточных мембран
Введение
Липиды являются не только структурными компонентами ЦНС, но и важнейшими участниками функциональной активности. Головной мозг характеризуется высоким содержанием липидов. Мозг содержит уникальные мембранные структуры – миелиновые оболочки, которые имеют самое высокое содержание липидов по сравнению с другими тканями или субклеточными структурами, за исключением адипозной ткани. Для ЦНС характерно и наибольшее структурное разнообразие липидов по сравнению с мембранами других органов.
Липидный состав нервной ткани практически постоянен и остается неизменным даже под влиянием внешних факторов, которые меняют липвдный состав висцеральных органов и плазмы. Это – следствие защищенности ЦНС от различных внешних воздействий. Изменение липидного состава нервной ткани рассматривается обычно как патология, хотя при этом следует помнить, что существенные изменения в липидном составе нервной системы происходят в период развития.
Вся сложнейшая деятельность нервной ткани опосредуется через мембраны, в формировании и функционировании которых липиды принимают непосредственное участие.
В клетках нервной системы представлено несколько типов высокоспециализированных мембран: соматические мембраны мульти- и униполярных нейронов, мембраны дендритов, миелинизированных и немиелинизированных аксонов, аксонного холмика, где генерируется потенциал действия, мембраны рыхлого и компактного миелина, мембраны синаптических пузырьков, пре- и постсинаптические мембраны, мембраны макро- и микроглии. Возбудимость этих мембран колеблется в широких пределах от высоковозбудимых до относительно устойчивых мультимембранных структур миелина. В составе, строении и функционировании мембран нервной ткани еще очень много неясного. Для того чтобы раскрыть надмолекулярную организацию этих мембран, надо иметь достаточно полное представление об их липидном и белковом составе. Однако исследователи пока не владеют этими сведениями в полной мере, хотя ряд важных закономерностей уже намечен.
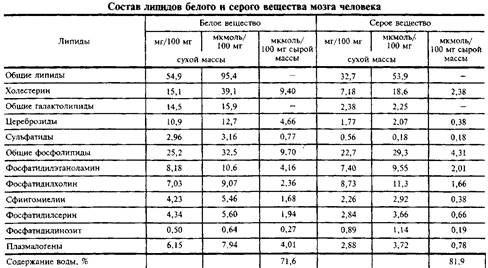
Липидный состав серого и белого вещества мозга человека представлен в табл. 1, а различных клеток мозга – в табл. 2. Видно, что липидный состав белого вещества ближе к миелину, а серое вещество содержит меньше типичных миелиновых ли-пидов, но относительно больше ганглиозидов.
Сравнивая молярное содержание основных классов липидов в специализированных клетках мозга, можно видеть, что оли-годендроглия и миелин наиболее обогащены цереброзидами, а нейроны и астроглия имеют более высокое содержание фосфолипидов. Это лишний раз подтверждает, что плазматические мембраны совершенно отличны от миелина.
Состав фосфолипидов обогащенных фракций нейронов и нейроглии коры мозга крысы представлен в табл. 3.
Чем более анатомически дифференцированно подходить к нервной ткани, тем больше различий обнаруживается в липидном составе, поскольку функционально различные нейрональ-ные и глиальные клетки имеют своеобразный липидный состав.
В состав большинства липидов входят жирные кислоты. В мозге они гораздо разнообразнее, чем в других тканях. Это намного увеличивает число индивидуальных липидов мозга. Содержание жирных кислот в головном мозге гораздо выше, чем в других органах, и составляет примерно 20–25% в расчете на сухую массу ткани. Разнообразие жирных кислот в этом органе поразительно. Применение газожидкостной хроматографии позволило продемонстрировать наличие в головном мозге более 50 жирных кислот с длиной цепи от 12 до 26 углеродных атомов, среди которых найдены насыщенные, ненасыщенные, нормальные, гидроксизамещенные, нечетные и др. Ненасыщенные кислоты мозга могут содержать от 1 до 6 двойных связей. Особенностью, липидов мозга является относительно большое содержание длинноцепочечных полиеновых кислот 20:4, 22: 5, 22:6.
Отдельные классы и фракции липидов мозга характеризуются своим набором жирных кислот. Имеет место также определенная специфичность жирнокислотного состава в лип идах разных отделов мозга, разных типов его клеток, субклеточных структур. Иллюстрацией этого могут служить данные табл. 4, где приведен жирнокислотный состав фосфолипидов синаптосом и миелина – двух разных типов мембранных структур ЦНС,
Состав липидов основных типов нервных клеток мозга крысы
| Липиды | Нейроны | Астроглия | Олигоден-дроглия | Миелин |
| Холестерин | 6,610 | 14,100 | 10,800 | 54,900 |
| Цереброзиды | 0,513 | 0,689 | 2,610 | 22,000 |
| Сульфатиды | 0,090 | 0,142 | 0,472 | 2,890 |
| Общие фосфолипиды | 22,400 | 35,600 | 23,400 | 41,800 |
| Ганглиозиды | 0,223 | 0,582 | 0,239 | 0,0453 |
| Молярное отношение: холестерин – цереброзиды-фосфолипиды | 1:0,075:3,5 | 1:0,05:2,5 | 1:0,25:2,2 | 1:0,40:0,76 |
| Фосфолипиды | Нейроны | Нейроглия |
| Лизофосфатидилхолин | 3,9 | 1,9 |
| Фосфатидилхолин | 46,1 | 46,9 |
| Сфингомиелин | 6,7 | 9,5 |
| Фосфатидилсерин | 9,1 | 7,1 |
| Фосфатидилинозит | 7,7 | 5,9 |
| Фосфатидил этанол а мин | 25,1 | 24,9 |
| Фосфатидная кислота | 1,8 | 3,6 |
Содержание индивидуальных фосфолипидов в коре мозга крысырезко различающихся по своему происхождению и функциям.
В синаптосомах велико содержание жирных кислот – С 22:6, а в миелине высок процент моноеновых кислот – 18:1. Возможно, что высокое содержание докозагексаеновой кислоты в синаптосомах необходимо для активного транспорта ионов, так как активность Na+ , К+ -АТФазы в них зависит от присутствия полиеновых кислот в фосфолипидах. В мозге имеются регуля-торные механизмы, поддерживающие степень ненасыщенности и специфичность жирнокислотного состава в липидах.
Состав жирных кислот фосфолипидов сннаптосомальных и миеляновых мембран коры мозга обезьяны
| Шифр жирной кислоты | Фосфатид ил- | |||||
| холи н | этанол амин | серин+монофосфо-инозитид | ||||
| синапто-сомы | миелин | синапто-сомы | миелин | синапто-сомы | миелин | |
| 16:0 | 50 | 33,1 | 7,4 | 4,9 | 3,5 | 2,6 |
| 18:0 | 12,4 | 17 | 25,5 | 15,9 | 44,3 | 43 |
| 18:1 | 27,2 | 42,3 | 12,1 | 33,2 | П, 4 | 38,5 |
| 20:1 | 0,7 | 0,9 | 1,6 ■ | 9,3 | - | 2,5 |
| 20:4 | 3,8 | 3,2 | 10,1 | 11,6 | 8,3 | 6,3 |
| 22:4 | 0,8 | 0,6 | 6,4 | 13,1 | 3,5 | 3,7 |
| 22:6 | 3 | 2,3 | 24,9 | 10,6 | 26,9 | 2,9 |
Изменение жирнокислотного состава приводит к нарушению функциональной деятельности мозга.
1. РОЛЬ АЦИЛОБМЕННОГО МЕХАНИЗМА
В мембранах головного мозга имеет место цикл деацилирование – реацилирование, при котором происходит замена жирных кислот в молекуле фосфолипидов, в то время как другие компоненты молекулы остаются неизменными. Этот ацилобменный механизм является особенно важным для включения тех или иных жирных кислот во второе положение остатка глицерина, и его рассматривают как средство локального регулирования физических и функциональных свойств мембран. Существенную роль играет и переход диацильных форм фосфолипидов в моноацильные и обратно. Все это оказывает влияние на такие мембранные процессы, как проницаемость для различных веществ, транспорт ионов и т.д.
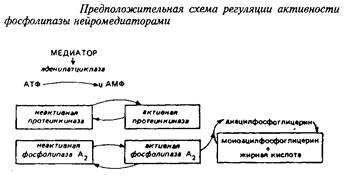
Ацилобменные реакции имеют прямое отношение ко многим процессам, влияя на активность ряда ферментов, на синтез простагландинов и чувствительность фоторецепторов. Некоторые исследователи связывают ферментативное деацилирование – реацилирование с эффектом синаптической передачи. Так, под влиянием норадреналина в синаптосомах происходит активирование фосфолипазы А2 , отщепляющей жирную кислоту во втором положении глицерофосфолипида. Таким образом, нейромедиатор модифицирует обмен фосфолигавдов в синаптических мембранах путем вовлечения в этот процесс реакций деацилирования. Предложена следующая схема регуляции активности ацилобменного цикла нейромедиаторами.
2. ОРГАНИЗАЦИЯ ЛИПИДОВ В МЕМБРАНЕ
Образование липидных молекул в ходе эволюции и выбор именно этих молекул в качестве строительных блоков мембран сыграли решающую роль в возникновении жизни. Липидам принадлежит жизненно важная роль в клетке. Следующие особые физико-химические свойства липидов определяют их роль в построении мембран:
1. Сочетание гидрофильных и липофильных свойств в структуре одной молекулы, их амфифильность.
2. Способность липидов четко ориентироваться на границе раздела фаз, так что полярные группы направлены в водные среды, а неполярные экранированы от них.
3. Способность липидов самопроизвольно упаковываться в прочные, плотные мономолекулярные слои или пленки, устойчивые к сжатию. Плотность такой упаковки зависит от рН, температуры и молекулярной организации липидов. Такие плотные слои создают определенный барьер для диффузии молекул.
4. Способность липидов агрегировать в хорошо упорядоченные сферические, цилиндрические, ламеллярные мицеллы. В мицеллах липиды ориентированы таким образом, чтобы максимальное число полярных групп находилось в контакте с водой, а гидрофобная часть была максимально удалена от контакта с ней.
Способность липидов образовывать прочные мономолекулярные слои лежит в основе молекулярной организации мембран. Более 60 лет назад было высказано предположение, что в основе мембран лежит бимолекулярный слой липидов.
В бимолекулярном липидном слое гидрофобные цепочки молекул липидов направлены друг к другу и внутренность бислоя совершенно гидрофобна, а гидрофильные части образуют поверхности внутреннего и внешнего монослоев, открытые для разнообразного рода взаимодействий.
--> ЧИТАТЬ ПОЛНОСТЬЮ <--