Курсовая работа: Механизмы проникновения вирусов в клетку. Биохимические и цитофизиологические аспекты
Слияние вирусной и клеточной мембран.
Для того чтобы внутренний компонент вируса мог пройти через клеточную мембрану, вирус использует механизм слияния мембран. У вирусов, имеющих суперкапсид, слияние обусловлено точечным взаимодействием вирусного белка слияния с липидами клеточной мембраны, в результате которого вирусная липопротеидная оболочка интегрирует с клеточной мембраной, а внутренний компонент вируса оказывается по другую ее сторону. У вирусов, не имеющих суперкапсид, один из поверхностных белков также взаимодействует с липидами клеточных мембран, в результате чего внутренний компонент проходит через мембрану. Большинство вирусов животных выходит в цитозоль из рецептосомы.
Если при эндоцитозе вирусная частица является «пассивным пассажиром», то при слиянии она становится активным участником процесса. Белком слияния является один из ее поверхностных белков (о них было упомянуто выше).
Общая схема проникновения вирусов с суперкапсидом в клетку:
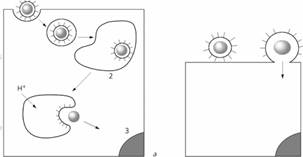
Слева — эндоцитоз и слияние в эндосоме: 1 — вирус, 2 — эндосома, 3 — ядро; справа— слияние с плазматической мембраной.
С позиций современной вирусной цитопатологии, классификация механизмов проникновения вирусов через плaзматическую мембрану пересмотрена в соответствии с открытием новых порталов для попадания питательных веществ в клетку. На данный момент выделяют пять регулируемых порталов: фагоцитоз, пиноцитоз, подразделяемый на макропиноцитоз и различные типы эндоцитоза - клатрин-опосредованный (размер образованной вакуоли ~120 нм), кавеолин-опосредованный (~60 нм), кавеолин- и клатрин-независимый эндоцитоз (~90 нм). Для вирусов присущи все вышеозначенные пути проникновения, поэтому необходимо более точное описание указанных порталов.
· Фагоцитоз у млекопитающих характерен для специализированных клеток моноцитов, макрофагов и нейтрофилов - и в этот процесс вовлекаются специфические рецепторы и последующий сигнальный каскад опосредуется определенными семействами ферментов - гуанинтрифосфатаз (ГТФазы). Данные ферменты играют ключевую роль для классификации порталов питательных веществ в клетки млекопитающих. Это большая группа ферментов, которые участвуют в связывании гуанинтрифосфатов и подвергаются конформационным изменениям при гидролизе гуанинтрифосфата в гуаниндифосфат в присутствии ионов Mg2+ . Наибольший интерес представляет семейство мономерных ГТФаз сигнальной каскадной трансдукции и движения (signal transduction cascades and motility, Ras), которые по химической структуре подразделяют на девять групп: Rho (А, В, С изоформы фермента), Rac (1, 2, 3 изоформы), Rab, Cdc42 (G25K, Cdc42Hs изоформы), RhoD, RhoG, TC10, Rnd (Rndl, RhoE/Rnd3, Rnd6) иTTF. Причем каждая группа участвует в определенных функциях клетки. Так, при фагоцитозе активируется Rhо-грyппа ГТФаз, связанная с актиновыми структурами цитоскелета, а ее инициация происходит при стимуляции рецепторов плазмалеммы. Rab-грyппа участвует в перемещении уже сформированных везикул в цитоплазме клеток. Таким образом, при фагоцитозе необходима стимуляция рецепторов клетки, после чего активируется Rhо-группа ГТФаз, приводящая в движение актиновые структуры цитоскелета.
· В сигнальный каскадный макропиноцитоз также вовлекается Rhо-группа ГТФаз, которая запускает актин-зависимое формирование выступов мембраны. В отличие от фагоцитоза выступы плазмалеммы не соединяются по принципу "молнии" с лигандами поверхности поглощенной частицы, а формируют большую эндоцитарную вакуоль. Мембрана этой вакуоли не связана с поверхностью поглощаемых частиц.
· Кавеолин-опосредованный эндоцитоз характеризуется образованием кавеол, которые формируются на специфичных участках плазмалеммы, где сконцентрированы разнообразные сигнальные молекулы, а также молекулы, ответственные за транспорт в плазмалемме клеток. Формирование этих структур связано с определенными липидными участками плазмалеммы в жидкой фазе, обогащенными холестеролом и ориентированными во внутрь мембраны сфингомиелинами и гликосфинголипидами. После интеграции белка кавеолина 1 с этим участком плазмалеммы, происходит инвагинация последней в форме небольшой фляжки, которая называется кавеолой. Кавеолин 1 - это мономер, который при стимуляции собирается в дискретный гомо-олигомер, включающий 14-16 кавеолиновых молекул. Смежные гомо-олигомеры упаковываются в пределах мембраны кавеолы, тем самым, обеспечивая структурную сеть для ее дальнейшего формирования. Причем поглощенный материал химически связан с рецепторами мембраны клетки.
· Следующий, клатрин-зависимый эндоцитоз прежде назывался как "рецептор-опосредованный". Данный вид эндоцитоза характеризуется концентрацией высокоаффинных трансмембранных рецепторов и связанных с ними лиганд в выемках плазматической мембраны. Покрытие этих выемок сформировано в процессе сборки специальных цитозольных белков, главным образом связанных с определенными единицами клатрина. Этот белок состоит из трех вытянутых тяжелых и плотно связанных цепей, собранных в полигональную упаковку.
При такой структуре клатрин способен деформировать плазмалемму, преобразуя ее поверхность в выемки. Эти выемки инвагинируют и формируют эндоцитарную вакуоль, изнутри покрытую полигонально упакованным клатрином. Таким образом сформированная эндоцитарная вакуоль переносит внутрь клетки комплекс трансмембранных рецепторов, соединенных с лигандами поглощаемого материала. При нефизиологической стимуляции клетки клатрин способен к спонтанной сборке в закрытую полигональную упаковку, тогда как при физиологических условиях для такой сборки полипротеина необходимо присутствие покрывающих выемку специальных белков (assemble proteins, АР) - мономерного АР180 и гетеротетрамерного протеинового комплекса адаптора APl-4, которые опосредуют формирование везикулы при ее субклеточной локализации. В последний комплекс входят две большие структурно связанные субъединицы, названные α- и β-адаптинами, центральная субъединица - µ2 и малая субъединица - σ2. Этот адаптивный комплекс назван "мускулом", который напрямую активирует клатрин к сборке для покрытия выемок, в результате чего изменяется цитоскелет и происходит инвагинация плазмалеммы. Место сборки клатрина определяет α-субъединица, привязывая комплекс АР2 к плазмалемме, тогда как остальные субъединицы привязаны специфично к другим органеллам клетки. В процессе клатрин-зависимого эндоцитоза принимают участие ГТФазы Arf- и Sаг-групп.
В последнее десятилетие в мембранах клеток млекопитающих исследователями были обнаружены микродомены, размером 40--50 нм, диффузно связанные с липидами. Эти участки преимущественно состоят из высокоорганизованных насыщенных липидов и холестерола, который латерально подвижен относительно второго уровня ненасыщенных липидов мембраны. При формировании инвагинаций плазмалеммы ограничение поглощенного материала в таких микроучастках происходит с помощью холестерола. Уникальный липидный состав участков, определяющих местоположение микродоменов, является физической основой для специфической сортировки белков мембраны, а гликолипиды составляют для них трансмембранную область.
· Кавеолин- и клатрин-независимый эндоцитоз. Исследования последних лет позволили отнести липидные участки клеточной мембраны к специфическому механизму эндоцитоза, в котором не используются кавеолин и клатрин. Описано несколько маркеров для этих липидных участков клеточной мембраны, такие как субблоки связанные с липидами бактериальных токсинов, а также рецепторы мембраны - гликосфинголипиды GM1, Gb3 и другие. С помощью ультрастуктурных и биохимических исследований установлен клатрин-независимый эндоцитоз интерлейкина 2 в лимфоцитах, который конститутивно возможен при наличии рецептора к этому цитокину. Это подтверждает известный факт, что лимфоциты лишены кавеолина и имеют рецептор к IL-2
Необходимо отметить роль динамина, который используется клетками при фагоцитозе, кавеолин- и клатрин-опосредованном эндоцитозах, а также в независимых от этих белков видах эндоцитоза. Этот протеин выступает как регулятор мембранного перемещения на поверхности клетки, является ГТФазой, молекулярной массой 100 кДа, и содержит несколько доменов, которые поддерживаются фосфотидилинозитол -4,5-бифосфатными связями. Информация о динамине закодирована в его собственном ГТФаз-активиру ющем белке внутри ГТФ-эффекторного домена. Впоздней стадии формирования везикул при взаимодействии ГТФазного эффекторного домена с другим концевым доменом – аргинином, динамин преобразуется в гексагональную кольцевую структуру типа «воротника», который отшнуровывает вакуоль от плазмалеммы. Тем не менее, точная функция динамина еще не определена, и на настоящий момент существуют 2 модели механизма его воздействия на отделение везикул от мембраны. Соответственно первой, динамин в отличие от остальных членов семейства ГТФаз может функционировать как механохимический фермент, при гидролизе ГТФ структурно сжимаясь. В этом случае динамин физически преобразуется в очень плотную структуру и как своеобразный жгут отшнуровывает везикулы от мембраны. Вторая, основана на наблюдениях, указывающих, что спираль, образованная динамином в присутствии ГТФ имеет большие промежутки. Это предполагает, что динамин действует как молекулярная пружина, проталкивая сформированную везикулу через цитозоль.
Как уже упоминалось выше, процесс перемещения вирусного генома и сопутствующих белков через мембрану клетки-хозяина называется пенетрацией или проникновением. Эволюция не создала для вирусов специфических структур, обеспечивающих их вход в клетку, и поэтому они используют клеточные рецепторы и порталы, существующие для проникновения жизненно-важных компонентов при нормальном функционировании клетки. Так, показано, что вирусы для входа в клетки-хозяева используют все эндоцитарные пути, включая клатрин-, кавеолин-опосредованные и недавно описанные независимые от этих белков виды эндоцитоза, а так же макропиноцитоз. Известны вирусы, которые могут проникать в клетки двумя механизмами, как с помощью эндоцитоза, зависимого от клатрина и кавеолина, так и путем, независимым от этих белков.
Для разграничения вирусов в зависимости от механизма проникновения в данной работе была использована вышеобозначенная классификация порталов для проникновения биологически активных веществв клетки (см. далее таблицу).Необходимо отметить, что некоторыми исследователями наряду с указанными механизмами входа вирусов также выделяются холестерол- и динамин-зависимые виды эндицитоза, но так как очень часто эти белки используются клеткой на различных этапах перемещения эндоцитарных везикул, то, возможно, нецелесообразно выделять последние два вида эндоцитоза.
Если рассматривать широкий спектр вирусов, то большинство из них для своего проникновения используют путь клатрин- опосредованного эндоцитоза (таблица). Данный путь физиологически наиболее эффективен, так как в этом случае вирусы защищены от воздействия на них биологически активных веществ цитоплазмы клеточной мембраной, формирующей эндоцитарную вакуоль. В пределах вакуоли вирусные частицы могут перемещаться и проникать в более глубокие области цитоплазмы с помощью механизма, получившего название "груз" - "cargo". Этот механизм осуществляется опосредовано через микротрубочки и используется для перемещения частиц размером больше 20 нм, так как из-за большого размера физические свойства цитоплазмы не позволяют самостоятельно везикулам свободно диффундировать. В этот процесс вовлекаются три различных класса биомолекул-моторов: кинезин, динеин и миозин.
| Семейства вирусов | Механизмы проникновения | ||
| "Неодетые" вирусы без суперкапсида | |||
| Пикорнавирусы | |||
| Полиовирус | Слияние вирусной оболочки и плазмалеммы клеток | ||
| и динамин-независимый эндоцитоз | |||
| Вирус ЕСН01 | Кавеолин-опосредованный эндоцитоз | ||
| род Пареховирусов, | Клатрин-опосредованный эндоцитоз | ||
| Риновирус человека 2 и 14 | Клатрин-опосредованный эндоцитоз | ||
| Риновирус человека 2 | Слияние вирусной оболочки и плазмалеммы клеток | ||
| Реовирусы | |||
| Ротавирус человека | Клатрин-опосредованный эндоцитоз и эндоцитоз, независимый | ||
| от клатрина и кавеолина | |||
| Паповавирусы | |||
| Вирус обезьян 40 | Кавеолин-опосредованный эндоцитоз | ||
| Вирус полиомы | Кавеолин-опосредованный эндоцитоз | ||
| Вирус папилломы коров | Слияние вирусной оболочки и плазмалеммы клеток | ||
| Вирус папилломы человека | Клатрин-опосредованный эндоцитоз | ||
| тип 16 и 58 | |||
| Парвовирусы | Клатрин-опосредованный эндоцитоз | ||
| Парвовирус псовых | Клатрин-опосредованный эндоцитоз | ||
| Минутный вирус мышей | Клатрин-опосредованный эндоцитоз | ||
| Аденовирусы | |||
| Аденовирус 5 | Клатрин-опосредованный эндоцитоз | ||
| Аденовирус 2 | Клатрин-опосредованный эндоцитоз и эндоцитоз независимый | ||
| от клатрина и кавеолина | |||
| Аденовирус 2 | Эндоцитоз независимый от клатрина и кавеолина | ||
| Аденовирус человека | Клатрин-опосредованный эндоцитоз и эндоцитоз независимый | ||
| от клатрина и кавеолина | |||
| Поксвирусы | |||
| Вакцинный вирус | Макропиноцитоз и слияние вирусной оболочки и плазмалеммы | ||
| клеток | |||
| Реовирусы | |||
| Инфекционные субвирусные | Клатрин-опосредованный эндоцитоз | ||
| частицы | |||
| "Одетые'' вирусы с наличием суперкапсида | |||
| Филовирусы | |||
| Вирус Марбурга | Кавеолин-опосредованный эндоцитоз | ||
| Вирус Эбола | Кавеолин-опосредованный эндоцитоз | ||
| Коронавирусы | |||
| Коронавирус человека | Кавеолин-опосредованный эндоцитоз | ||
| Алъфавирусы | |||
| Вирус леса Семлики | Клатрин-опосредованный эндоцитоз | ||
| Синбис вирус | Клатрин-опосредованный эндоцитоз | ||
| Флавивирусы | |||
| Вирус клещевого энцефалита | Клатрин-опосредованный эндоцитоз | ||
| Вирус японского энцефалита | Клатрин-опосредованный эндоцитоз | ||
| Вирус западного Нила | Клатрин-опосредованный эндоцитоз | ||
| Семейства вирусов | Механизмы проникновения | ||
| Ортомиксовирусы | |||
| Вирус гриппа А | Клатрин-опосредованный эндоцитоз и клатрин-независимый эндоцитоз | ||
| Рабдовирусы | |||
| Вирус везикулярного стоматита | Клатрин-опосредованный эндоцитоз | ||
| Буньявирусы | |||
| Вирус La Crosse | Клатрин-опосредованный эндоцитоз | ||
| Хантавирус | Клатрин-опосредованный эндоцитоз | ||
| Иридовирусы | |||
| Вирус африканской лихорадки свиней | Клатрин-опосредованный эндоцитоз | ||
| Тогавирусы | |||
| Вирус краснухи | Клатрин-опосредованный эндоцитоз | ||
| Парамиксовирусы | |||
| Вирус мозаики | Слияние вирусной оболочки и плазмалеммы клеток | ||
| Ортогепаднавирусы | |||
| Вирус черного гепатита В | Клатрин-опосредованный эндоцитоз | ||
| Вирус гепатита мыши 4 | Клатрин-опосредованный эндоцитоз и слияние вирусной оболочки и плазмалеммы клеток | ||
| Ретровирусы | |||
| Вирус иммунодефицита человека | Макропиноцитоз и слияние вирусной оболочки и плазмалеммы клеток | ||
| Вирус лейкоза | Клатрин-опосредованный эндоцитоз | ||
| Герпесвирусы | |||
| Вирус простого герпеса 1 | Слияние вирусной оболочки и плазмалеммы клеток | ||
| Вирус простого герпеса 1 | Клатрин-опосредованный эндоцитоз | ||
| Вирус герпеса 4 Эпстайна-Барр | Слияние вирусной оболочки и плазмалеммы клеток и клатрин-опосредованный эндоцитоз | ||
| Вирус герпеса 1 и 5 у коров | Слияние вирусной оболочки | ||
| Цитомегаловирус | Клатрин-опосредованный эндоцитоз | ||
| Вирус герпеса 8 (вирус саркомы Капоши) | Клатрин-опосредованный эндоцитоз | ||
| Бакуловирусы | Клатрин-опосредованный эндоцитоз и макропиноцитоз | ||
| Артеривирусы | |||
| Вирус респиративного синдрома у свиней | Клатрин-опосредованный эндоцитоз | ||
Принято, что для "одетых" вирусов с наличием суперкапсида проникновение осуществляется путем слияния их оболочки с плазмалеммой клетки либо путем рецептор-опосредованного эндоцитоза. Последний используют вирус леса Семлински и вирус гриппа А. Механизм прямого слияния с плазмалеммой характерен для вируса Сен-дай и некоторых ретровирусов. Различие механизмов слияния клеточной мембраны с суперкапсидом вирусов определяется наличием у них интегральных гликопротеинов - белков слияния. Они отличаются способом активации и структурными особенностями, и их функционирование зависит от присутствия трансмембранных гликопротеинов.
Исследование структур и функций белков слияния позволило разделить их на два класса по радиально отличающейся архитектонике. Конформационные изменения эктодоменов этих белков обеспечивают процесс слияния с мембраной клетки. Предполагается, что расширенный трехнитевой стержень белков является α-спиральным и имеет сходство с общей структурой и функциями эктодоменов белков слияния различных вирусов, имеющих суперкапсид. Таким образом, отмечаются общие черты процесса слияния вирусной оболочки с плазмалеммой при проникновении различных вирусов этой группы в клетки. В этом случае при активации стабильный белок слияния пространственно изменяется и приобретает стержневидную форму, подобную "шпильке", которая внедряется в определенный участок клеточной мембраны. Затем обратная сторона белка слияния складывается на начало, тем самым, заключая белок слияния и трансмембранную область в себя, что приводит к близкому контакту вирусной оболочки и мембраны. В этом процессе в основном участвуют липиды плазмалеммы. В дальнейшем, происходит образование локальной полусферы с последующим открытием и увеличением поры в клеточной мембране для передачи вирусных компонентов.
Недавние исследования показали, что при последней стадии слияния суперкапсида вирусов с плазмалеммой, т.е. при формировании и увеличении поры слияния, необходимы определенные клеточные трансмембранные области минимальной длины. При этом не определена временная зависимость процесса формирования поры слияния от точной последовательности химической структуры подобной трансмембранной области, но есть необходимость в ее определенных функциональных свойствах. Такие липидные участки плазмалеммы, используемые вирусами как платформы для входа, обеспечены локализованной концентрацией рецепторов и/или сопутствующими им корецепторными комплексами, а также другими компонентами клетки, которые модулируют процесс входа. Эти участки имеют определенное значение для завершения процесса слияния суперкапсида вируса с плазмалеммой, особенно физические свойства липидов, составляющие участок контакта с оболочкой вируса. При этом также имеет определенное значение структурный состав вирусного суперкапсида. Так, в случае, если в нем преобладают липиды, для входа вируса отпадает необходимость в трансмембранных участках плазмалеммы. Указываются некоторые исключения, когда при проникновении вируса в клетки при функционировании вирусных белков слияния не используются его внутренние домены.
Исследования последних лет ?