Курсовая работа: рДНК-биотехнология. Способы биотрансформации клеток
Третий этап - начало работ по включению в векторные молекулы ДНК (ДНК, используемые для переноса генов и способные встраиваться в генетический аппарат клетки-рецепиента) генов эукариот, главным образом, животных.
Формально датой рождения генетической инженерии следует считать 1972 год, когда в Стенфордском университете П. Берг, С. Коэн, Х. Бойер с сотрудниками создали первую рекомбинантную ДНК, содержавшую фрагменты ДНК вируса SV40, бактериофага и E. coli.
1 Основные группы ферментов генетической инженерии
Генетическая инженерия - потомок молекулярной генетики, но своим рождением обязана успехам генетической энзимологии и химии нуклеиновых кислот, так как инструментами молекулярного манипулирования являются ферменты. Если с клетками и клеточными органеллами мы подчас можем работать микроманипуляторами, то никакие, даже самые мелкие микрохирургические инструменты не помогут при работе с макромолекулами ДНК и РНК. Что же делать? В роли "скальпеля", "ножниц" и "ниток для сшивания" выступают ферменты.
Только они могут найти определенные последовательности нуклеотидов, "разрезать" там молекулу или, наоборот, "заштопать" дырку в цепи ДНК. Эти ферменты издавна работают в клетке, выполняя работы по репликации (удвоению) ДНК при делении клетки, репарации повреждений (восстановлению целостности молекулы), в процессах считывания и переноса генетической информации из клетки в клетку или в пределах клетки. Задача генного инженера - подобрать фермент, который выполнил бы поставленные задачи, то есть смог бы работать с определенным участком нуклеиновой кислоты.
Следует отметить, что ферменты, применяемые в генной инженерии, лишены видовой специфичности, поэтому экспериментатор может сочетать в единое целое фрагменты ДНК любого происхождения в избранной им последовательности. Это позволяет генной инженерии преодолевать установленные природой видовые барьеры и осуществлять межвидовое скрещивание.
Ферменты, применяемые при конструировании рекомбинантных ДНК, можно разделить на несколько групп:
- ферменты, с помощью которых получают фрагменты ДНК (рестриктазы);
- ферменты, синтезирующие ДНК на матрице ДНК (полимеразы) или РНК (обратные транскриптазы);
- ферменты, соединяющие фрагменты ДНК (лигазы);
- ферменты, позволяющие осуществить изменение структуры концов фрагментов ДНК.
1.1 Рестриктазы
Общепринято термины "рестриктаза", "эндонуклеаза рестрикции" и "сайт специфическая эндодезоксирибонуклеаза" считать синонимами.
Все рестрикционные эндонуклеазы бактерий узнают специфические, довольно короткие последовательности ДНК и связываются с ними. Этот процесс сопровождается разрезанием молекулы ДНК либо в самом сайте узнавания, либо в каком-то другом, что определяется типом фермента. Наряду с рестрикционной активностью бактериальный штамм обладает способностью метилировать ДНК; для этого процесса характерна такая же специфичность в отношении последовательностей ДНК, как и для рестрикции. Метилаза добавляет метильные группы к адениновым или цитозиновым остаткам в том же сайте, в котором связывается рестрикционный фермент. В результате метилирования сайт становится устойчивым к рестрикции. Следовательно, метилирование защищает ДНК от разрезания.
Различают 3 основных класса рестриктаз: 1, 2 и 3.
Все рестриктазы узнают на двуспиральной ДНК строго определенные последовательности, но рестриктазы 1-го класса осуществляют разрывы в произвольных точках молекулы ДНК, а рестриктазы 2-го и 3-го классов узнают и расщепляют ДНК в строго определенных точках внутри сайтов узнавания или на фиксированном от них расстоянии.
Ферменты типов 1 и 3 имеют сложную субъединичную структуру и обладают двумя типами активностей - модифицирующей (метилирующей) и АТФ-зависимой эндонуклеазной.
Ферменты второго класса состоят из 2 отдельных белков: рестрицирующей эндонуклеазы и модифицирующей метилазы, поэтому в генной инженерии используются исключительно ферменты 2-го класса. Они нуждаются в ионах магния в качестве кофакторов.
В настоящее время выделено более 500 рестриктаз класса 2, однако среди ферментов, выделенных из различных микроорганизмов, встречаются такие, которые узнают на ДНК одни и те же последовательности. Такие пары или группы называют изошизомерами. Различают истинную изошизомерию, когда ферменты узнают одну и ту же последовательность нуклеотидов и разрывают ДНК в одних и тех же точках, и ложную, когда ферменты, узнавая один и тот же сайт на ДНК, производят разрывы в разных точках в пределах того же сайта.
Большинство рестриктаз класса 2 узнают последовательности, содержащие от 4 до 6 нуклеотидных пар, поэтому рестриктазы делят на мелко- и крупнощепящие. Мелкощепящие рестриктазы узнают тетрануклеотид и вносят в молекулы гораздо больше разрывов, чем крупнощепящие, узнающие последовательность из шести нуклеотидных пар. Это связано с тем, что вероятность встречаемости определенной последовательности из четырех нуклеотидов гораздо выше, чем последовательности из шести нуклеотидов. Например, в ДНК бактериофага Т7, состоящей из 40000 пар оснований, отсутствует последовательность, узнаваемая рестриктазой R1 из E. coli.
К мелкощепящим относятся рестриктазы Hpa II и Alu (из Arthrobacter luteus), к крупнощепящим - Eco R I (из Escherichia coli) и Hind III. Если предположить, что участки узнавания рестриктаз распределены вдоль цепи ДНК случайно, то мишень для ферментов, узнающих последовательность (сайт) из четырех нуклеотидов, должна встречаться в среднем 1 раз через каждые 256 пар оснований, а для ферментов, узнающих шесть нуклеотидов, - через 4096 пар оснований. Если сайт рестрикции окажется внутри гена, то обработка ДНК-рестриктазой приведет к его инактивации. Вероятность такого события очень велика при обработке мелкощепящими рестриктазами и незначительна при применении крупнощепящих эндонуклеаз. Поэтому с целью получения неповрежденного гена расщепление проводят поочередно несколькими крупнощепящими рестриктазами, либо применяют прием "недорестрикции", т.е. рестрикцию проводят в таких условиях, когда происходит расщепление лишь в одном сайте.
1.1.1 Механизм действия рестриктаз
В качестве мишеней (мест узнавания) часто выступают палиндромы из 4-6 пар оснований - сайты рестрикции. Точки узнавания рестриктазами симметричны относительно поворота на 180оС, то есть последовательность нуклеотидов слева направо в одной нити такая же, как справа налево в другой. Симметрия подразумевает, что те из них, которые должны быть метилированы, встречаются на обеих цепях ДНК. В результате сайт-мишень может быть полностью метилирован (обе цепи модифицированы), полуметилирован (только одна цепь метилирована) или не метилирован.
Полностью метилированный сайт не подвержен ни рестрикции, ни модификации. Полуметилированный сайт не узнается ферментом рестрикции, но может быть превращен с помощью метилазы в полностью метилированный. У бактерий метилирование, как правило, связано с сохранением имеющегося состояния модификации. Репликация полностью метилированной ДНК ведет к образованию полуметилированной ДНК. Вероятно узнавание полуметилированных сайтов представляет собой обычный этап функционирования метилазы in vivo.
Неметилированный сайт-мишень пред¬ставляет собой субстрат либо для рестрикции, либо для модификации in vitro. В клетке немодифицированная ДНК с большей вероятностью рестрицируется. Реакция разрезания осуществляется в две ступени. Сначала разрезается одна цепь ДНК, а затем рядом разрезается другая. В областях, прилегающих с каждой стооны к сайту разрезания, может иметь место экзонуклеотическая деградация. Происходит эффективный гидролиз АТФ, роль которого еще не выяснена.
Каким образом фермент узнает один сайт, а разрезает другой, достаточно удаленный? Важно отметить, что белок никогда не отделяется от молекулы ДНК, с которой он первоначально связался. Если фермент инкубировать со смесью модифицированной и немодифицированной ДНК, он предпочтительно разрезает немодифицированную ДНК. Следовательно, узнавая сайт связывания, белок не отделяется от неметилированной ДНК для того, чтобы найти сайт разрезания.
Существуют две альтернативные модели, объясняющие взаимосвязь между сайтами узнавания и разрезания: в соответствии с одной из них движется фермент, согласно другой модели, перемещается ДНК. Если движется фермент, то его перемещение вдоль ДНК будет продолжаться до тех пор, пока он не сделает выбор сайта разрезания. Если же движется ДНК, то фермент остается прикрепленным в сайте узнавания, а ДНК протаскивается через второй сайт связывания на ферменте, и это продолжается до тех пор, пока фермент не достигает области разрезания (пока не охарактеризованной). Получены электронно-микроскопические данные, свидетельствующие, что фермент вызывает образование петли в ДНК и остается, по-видимому, связанным с сайтом узнавания после разрезания; эти данные подтверждают вторую модель.
1.1.2 Построение рестрикционных карт
Ферменты рестрикции стали эффективным инструментом исследования. Они позволяют превращать молекулы ДНК очень большого размера в набор фрагментов длиной от нескольких сотен до нескольких тысяч оснований. С помощью метода электрофореза в агарозном геле (см. раздел 1) фрагменты ДНК, различающиеся по размеру, можно легко разделить, а затем исследовать каждый фрагмент отдельно.
Короткие фрагменты мигрируют намного быстрее, чем длинные. При сравнительно высокой концентрации агарозы большие фрагменты вообще не могут проникнуть в гель. В процессе миграции рестрикционные фрагменты не деградируют, их можно элюировать (вымывать) в виде биологически активных двуцепочечных молекул. При окрашивании гелей красителями, связывающимися с ДНК, выявляется набор полос, каждая из которых отвечает рестрикционному фрагменту, молекулярную массу которого можно определить, проведя калибровку с помощью ДНК с известными молекулярными массами.
Сравнение размеров фрагментов ДНК, полученных после обработки определенного участка генома набором рестрицирующих нуклеаз позволяет построить рестрикционную карту, на которой указано положение каждого сайта рестрикции относительно других участков.
Молекулу ДНК длиной 5000 пар нуклеотидов (п. н.). обрабатывают отдельно рестриктазами А и В. Фрагменты разделяют электрофорезом. Фермент А разрезал ДНК на 4 фрагмента размером 2100, 1400, 1000 и 500 п. н. Обработка рестриктазой В дала 3 фрагмента: 2500, 1300 и 1200 п. н. (рис. 37). Для определения расположения сайтов рестрикции этих ферментов на следующем этапе применяют процедуру двойного расщепления – обрабатывают ДНК двумя эндонуклеазами. Обработка изучаемого фрагмента одновременно двумя рестриктазами дала 6 фрагментов: 1900, 1000, 800, 600, 500, 200 п. н.
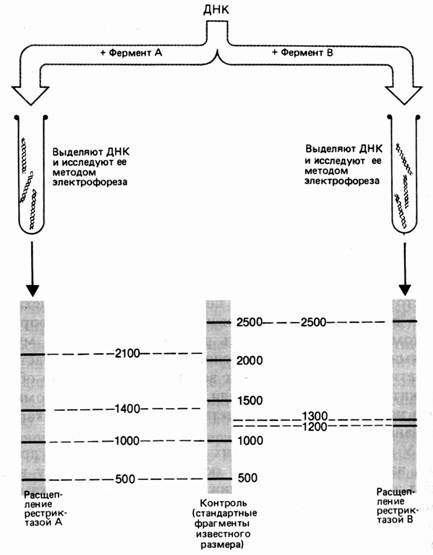
Рис. 1. Результаты электрофореза после обработки фрагмента ДНК разными рестриктазами
Наиболее полный вариант – элюировать каждый фрагмент, образующийся в результате расщепления одной рестриктазой, а затем обработать его второй. Смесь фрагментов, полученных после такой обработки, также анализируют с помощью электрофореза. В нашем примере были получены следующие результаты: