Курсовая работа: Розвиток кісткової тканини в онтогенезі безхвостих земноводних
Останні через відповідні канали проникають у кісткову тканину та кістковий мозок. Кісткові пластинки компактної речовини діафізів трубчастих кісток утворюють зовнішню загальну систему пластинок, остеонний шар, вставні системи і внутрішню загальну систему кісткових пластинок. Кісткові пластинки зовнішньої загальної системи розміщені паралельно до поверхні кістки і тісно зв'язані з окістям. Остеонний шар займає більшу площу кістки.
Остеон — сукупність 4— 20 трубчастих пластинок, вставлених одна в другу; за своєю формою вони мають вигляд спиляного дерева. У копитних тварин остеони побудовані з пластинок з майже горизонтальним або вертикальним спрямуванням колагенових волокон, що забезпечує міцність та пружність. їх центральні канали містять судини, остеобласти та остеокласти. Між остеонами знаходяться вставні пластинки, що являють собою залишки остеонів у стані моделювання.
Внутрішні загальні пластинки межують з ендоостом, останній відмежовує кістковомозкову порожнину і складається з тонковолокнистої сполучної тканини і містить остеобласти та остеокласти. В пластинках зовнішньої загальної системи проходять проривні пучки і канали з кровоносними судинами та нервами, розгалужуються, з'єднуються з центральними каналами остеонів і анастомозують між собою. Остеоцити лежать між кістковими пластинками, а кісткові канальці пронизують системи пластинок.
Розвиток кісткової тканини — остеогенез. Існує два різновиди остеогенезу — безпосередньо із мезенхіми та заміни кісткою гіалінового хряща. Ділянки мезенхіми, в яких почався остеогенез за зовнішнім виглядом являють собою мембрани, осифікація у них називається інтрамембранною. Цей остеогенез характерний для розвитку кісток черепа та нижньої щелепи, на місці, де утворюється кістка, спочатку знаходиться шар рихлої мезенхіми, вона має вигляд зірчастих клітин з відростками, які вступають в контакт з відростками сусідніх клітин мезенхіми, утворюючи сітку, остання занурена в аморфну міжклітинну речовину, що містить окремі пучки колагенових волокон і кровоносні судини. Окремі мезенхімні клітини відмежовані міжклітинною речовиною. На поверхні таких остеогенних острівців, швидко і непомітно переходять через стадію остеогенних клітин, диференціюються в остеобласти, що беруть участь в остеогенезі. Окремі остеобласти втрачають здатність синтезувати міжклітинну речовину, замуровуються в неї і перетворюються в остеоцити, міжклітинна речовина насичується фосфатом кальцію, який нагромаджується в кістковій тканині у результаті розпаду гліцерофосфату крові під впливом лужної фосфатази. Фосфат кальцію та вуглекислий кальцій, що при цьому утворюються, насичують основну речовину кісткової тканини. В зоні мінералізації остеобластами інтенсивно секретуються глікозаміноглікани, які потім розщеплюються лізосомними ферментами, утворюючи при цьому високоактивні аніони. В цьому процесі значне місце належить АТФ, яка може бути одночасно і джерелом фосфору, необхідного для кальцифікації й джерелом енергії. Цей тип остеогенезу характерний для розвитку первинної грубоволокнистої кісткової тканини. В подальшому первинна грубоволокниста тканина заміщується пластинчастою кістковою тканиною.
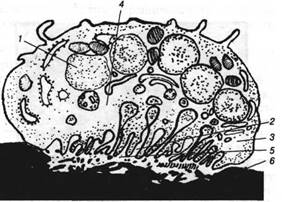
Рис. 3. Схема будови остеокласта:
1 — ядро; 2 — гофрований край остеокласта; 3 — світла зона; А — лізосоми; 5 — зона резорбції міжклітинної речовини; 6 — мінералізована міжклітинна речовина
Остеогенез на місці хряща. Інший різновид остеогенезу характерний для кісток тулуба, кінцівок, основи черепа. Він також відбувається в мезенхімі, однак йому передує формування моделі з гіалінового хряща, який руйнується і замінюється кістковою тканиною. При цьому в ділянці навколо хряща виникає волокниста кісткова манжетка, що сприяє появі перихондрального окостеніння. Воно відбувається за рахунок міграції із судин охрястя остеогенних клітин, які продукують осеїнові волокна й хондромукоїд, відбувається їх звапнування. Поступово манжетка потовщується і росте в напрямку епіфізів. Вона відмежовує ріст хряща в товщину і зумовлює формування у ньому клітинних колонок. Клітини хряща збільшуються в розмірі, набувають пузирчастого вигляду, їх ядра зморщуються, розпадаються. Міжклітинна речовина хряща між колонками клітин ущільнюється, заповнюється, утворює хрящові балки. Настає енхондральне окостеніння, яке супроводжується утворенням діафізарного центру окостеніння, що починається з проростання кровоносних судин кісткової манжетки всередину діафізу хрящової моделі; з судинами мігрують різнодиференційовані мехенхімні клітини гемопоетичного ряду й остеогенні. За рахунок остеокластів у хрящі виникають порожнини резорбції. Останні з'єднуються і утворюють кістково-мозкову порожнину. Малодиференційована сполучна тканина утворює первинний кіст-ковий мозок, збагачений остеобластами та малодиференційованими елементами, які в подальшому перетворюються у елементи дефінітивного кісткового мозку.
Навколо кровоносних судин утворюються кісткові пластинки, формуються остеони. В подальшому енхондральна кісткова тканина повністю розсмоктується і на її місці утворюється кістково-мозковий канал, зникає примітивна грубоволокниста кісткова тканина. Одночасно з енхондральним остеогенезом з боку окістя відбувається активний перихондральний остеогенез. Періостальна кісткова тканина являє собою компактну речовину кісток скелету, її будова типова пластинчастій з характерними системами кісткових пластинок. В епіфізах кісток центри окостеніння виникають пізніше. Окостеніння, яке виникає у діафізі, поширюється в напрямку епіфізів. У них з'являються місця окостеніння, за винятком хряща, що покриває суглобову поверхню та хрящову пластинку росту (епіфізарну лінію), яка відмежовує епіфіз від діафізу. Хрящ суглобової поверхні зберігається протягом усього життя тварини; у дорослих тварин хрящ епіфізарної лінії костеніє. За рахунок хряща епіфізарної лінії кістка росте в довжину.
Регенерує кісткова тканина за рахунок камбіальних елементів окістя. Перебудова кісток відбувається протягом усього життя тварини.
Розділ 2. Особливості будови окістя в безхвостих амфібій
На стегнових кістах амфібій (личинки Rana temporarіa, Pelobates fuskus, Hyla arborea на стадії 46, 47, 49 по Терентьєву, 1950; на стадії метаморфоза личинки, сеголіток і статевозрілих амфібій) у різні періоди онтогенезу вивчалися початкові стадії появи періосту і його наступний розвиток у процесі заміщення хрящової закладки кістою зі звертанням особливої уваги на клітинний склад і загальну архітектоніку за морфологічними і гістохімічними ознаками.
Шматочки стегнової кісти разом з періостом або тільки відщиплений періост вирізували із середньої частини діафізу, зони проксимального і дистального метафізів.
Відібраний матеріал фіксували в рідині Карнуа, 10 %-м нейтральному формаліні, 6 %-м формаліні, на насиченому розчині бензойної кислоти, у 80° і 96°-м етанолі при кімнатній температурі і при +4°С. У тих випадках, коли періост вирізували з підлягаючою кістою, відібрані шматочки поміщали в 10 %-й розчин динатриєвої солі етилендіаментетрауксусной кислоти при 18 °С (рН 7,4). Підготовлені зразки заливали в парафін. Гістологічні зрізи (5-6 мкм) офарблювали гематоксилин-еозином по Майеру і по Делафильду, а також селективними методами для виявлення визначених структур; у клітках визначали мітотичний індекс площі кліток, їхній ядерно-плазменне відношення. Числові показники оброблені методами варіаційної статистики.
У личинок на стадії 46 періост починає диференціюватися в діафізарній зоні майбутньої кісти. Перші ознаки периостальних структур з'являються на основі перихондра ще до виникнення тут кісткової пластинки. У цей час у найглибшому шарі перихондра клітини набувають характерного вигляду остеобластів і починають продукувати фермент лужну фосфатазу. Ці ознаки свідчать про початок розгортання периостального остеопластичного процесу. Очевидно, поява перших остеобластів зв'язано з проникненням кровоносних судин у перихондр і остеогенного диференціювання клітин перихондра в локусах, підготовлених до периостальному остеогенезі. Діяльністю цих клітин по окружності діафізарної хрящової закладки у вигляді такого пояса починає формуватися периостальна кісткова манжетка.
Клітини, що мають властивості остеобластів, морфологічно виділяються в новому періості більш великими розмірами, округлим ядром із сітчастою структурою і масивною цитоплазмою зі слабкими оксифільними властивостями. Для них характерна також визначена топографія: вони розташовуються безпосередньо в поверхні закладки і повернені до неї цитоплазматичной масою, тоді як ядро в кожній клітині знаходиться на протилежному кінці. Такий полярний розподіл ядра і цитоплазми додає зрілому функціонуючому остеобластові своєрідний вид комети, поверненої "хвостом" до споруджуваної кісти (мал. 4). Остеобласти, що дозрівають, (преостеобласти) мають більш симетричні контури з центральною локалізацією ядра. Маси цитоплазми в них не перевищує 1/3 її обсягу в зрілих формах. По фронті остеогенеза остеобласти в періості розташовуються в один ряд. Тут майже немає волокнистих структур, фіброзний каркас розташовується трохи вище і зв'язаний безпосередньо з преостеобластичним диференціюванням. Загальна товщина періосту в діафізарній зоні не перевищує 52 мкм.
У метафізарній зоні трубчастої кісти вже на 46-й стадії остеогенезу жаби мається типове для земноводних грибоподібне розростання епіфізарного хряща (мал. 5). У круговій щілині між диафизарной областю кісти, що розвивається, і хрящовим дивертикулом її епіфіза з моменту появи діафізарної кісткової манжетки сусідять на невеликій відстані кіста і хрящ. У цій області зберігається також своєрідна структура, що свідчить не тільки про можливу функціональну сумісність перихондра і періосту, але і про їхню єдність по генезу.
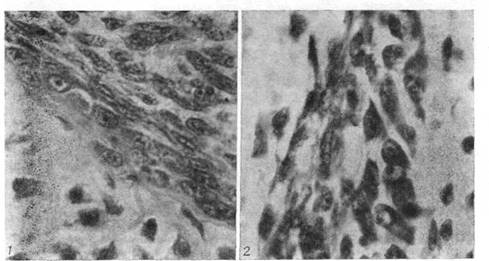
Рис. 4. Перихондр закладки стегнової кістки Rana temporarіa (в області диафиза). Видні остеобласти, що диференціюються, у внутрішньому шарі перихондра (гематоксилін Делафильда - эозин; про. 40, ок. 7).
Рис. 5. Періост області завороту хрящового епіфіза і метафиза стегнової кістки жаби. Між хрящовим епіфізом і кісткою розташовуються скелетогенные клітини (гематоксилін Делафильда - еозин; про. 60; ок. 7).
Перихондрально-періостальна ділянка включає всього 2-3 ряду клітин, з яких внутрішній шар складається з остеобластів, а зовнішній - з хондробластів (мал. 5). У кінцевому верхівковому відділі щілини, куди край кісткової трубки не досягає, утримуються тільки хондробласти.
У личинок 47-й стадії розвитку в трубчастих кістах кінцівок зберігається та ж ситуація, що описана для стадії 46 з тією лише різницею, що перихондральна кісткова трубка в області діафізу має трохи більші розміри по довжині і масі. Якщо на попередній стадії зачаток кісткового діафізу мав вигляд на гістологічному зрізі як тонка (10-12 мкм) пластинка, то на стадії 47 вона досягає в товщину 24 мкм і вже містить у своєму складі одиничні остеоцити (включені в кісту остеобласти) (мал. 6). У центральній частині діафізу остеобласти періосту здобувають подовжену форму і подовжнє розташування без вираженого полярного орієнтування до фронту остеогенеза (мал. 6). Зміна форми в остеобластах, як правило, сполучено з їхньою функціональною активністю (Мажуга, 1978; Вечерська, 1984).
Можна думати тому, що в серединній зоні діафіза кістки, що розвивається, остеопластичний процес здобуває поступово меншу інтенсивність, чим у метафізах.
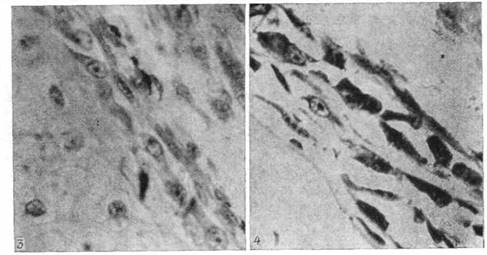
Рис. 6. Періост центра диафиза стегнової кісти R. temporarіa на 47-й стадії розвитку. Відзначаються одиничні остеоцити в периостальній кістці (гематоксилін Майера - еозин; про. 40, ок. 7).
Рис. 7. Періост зони діафіза кісти жаби на 49-й стадії розвитку. Відзначається вогнище енхондральної резорбції, у якому вже помітні зачатки миєлогенного ретикулума (гематоксилін - еозин; про. 40, ок. 7).
Клітини другого (зовнішнього) ряду морфологічно більш різнорідні. Серед них зустрічаються преостеобластические стадії з порівняно великим ядром. І ті й інші зв'язані з коллагеновым каркасом, утвореним, очевидно, їхньою участю.
Кількісний розподіл кліток у різних зонах періосту характеризується наступними показниками: у діафізарної області протягом 340 мкм утримується 11 остеобластів і 10 фібробластоподібних кліток.
У личинок трав'яної жаби 49-й стадії розвитку в стегновій кісті помітне насамперед збільшення в її розмірах по довжині й у поперечнику. Перихондральна кісткова трубка добре сформована. У центрі діафіза в хрящовому стрижні до цього часу з'явилося вогнище енхондральної резорбції, у якому вже помітні зачатки мієлогенного ретикулума (мал. 7). У проксимальному і дистальному відділах хрящового стрижня чітко виступає зональна диференціювання хондроцитів, у зв'язку з чим по довжині хрящового стрижня виділяються території: епіфізів, пролиферируючих хондроцитів, гіпертрофованих хондроцитів, що відрізняються своєї цитоархітектонікой.
Періост також здобуває в масі переважно за рахунок збільшення кількості остеобластів, що розташовуються, як правило, у два ряди, і фібробластоподобних клітин. Цей показник відповідає помітної інтенсифікації остеопластичного процесу в зв'язку з прискоренням темпів росту кісткової ланки і кістяка в цілому. Товщина періосту зберігається більш-менш рівномірної на всьому протязі і складає приблизно 50 мкм.