Курсовая работа: Розвиток кісткової тканини в онтогенезі безхвостих земноводних
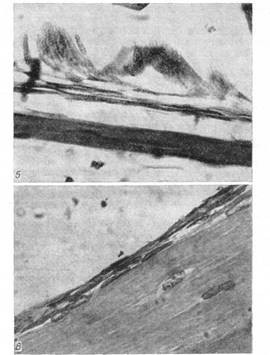
Рис. 8. Періост проксимального метафиза стегнової кісти жаби після завершення метаморфоза. Видні подовжені за формою остеобласти (гематоксилін Майера - эозин; про. 40, ок. 7).
Рис. 9. У сформованій стегновій кісті статевозрілої R. temporarіa окістя представлене одношаровою структурою (гематоксилін Майера - эозин; про. 90, ок. 7).
Природно, виникає питання: яким шляхом поповнюється склад остеогенних клітин у періості? На гістологічних зрізах довгих кіст трав'яної жаби личинокових стадій розвитку в періості ми завжди знаходили клітки в стані мітозу. Частка цих кліток невелика: серед остеобластів мітотичний індекс не перевищує 0,2 %, а для кліток менш диференційованих (у зовнішньому шарі) він досягає 1,0-1,5 %. З цих спостережень можна припустити, що періост самопідтримується тканинними системам. Поповнення кліток у ньому відбувається переважно за рахунок менш диференційованих станів і частково за рахунок функціонально зрілих остеобластів.
З початком метаморфоза, коли стегнова кіста в зростаючій кінцівці вже майже сформована, періост у діафізарної області переходить у зрілий стан і його клітки, включаючи остеобласти, здобувають компактну-подовжену форму з порівняно дрібним щільним ядром також подовженої форми. У структурі періосту до цього часу підсилюється колагеновий волокнистий каркас, і весь періост стає більш щільним і міцним. Зрілий періост можна тому, додаючи визначене зусилля, відокремити від кісти у виді тонкої плівки, чого не можна зробити з періостом тільки формується, при такій спробі він звичайно руйнується.
У кістках, що закінчили ріст, або в періоди сезонного припинення росту клітки періосту стають морфологічно однотипними (мал. 9). Серед них не зустрічаються з ознаками функціонуючих остеобластів, хоча всі клітки такого періосту є потенційно остеогенними. При зміні умов у ньому можуть активізуватися остеопластичні функції з появою морфологічно ідентифікованих остеобластів у зонах поновлення процесів ремоделювання кістки або її росту.
З цієї причини в дорослих жаб у період літнього активного стану завжди виявляються зональні особливості в структурі періосту, кількості в ньому клітинних рядів, а також формі і розмірах самих клітин. При цьому в діафізарної області структура періосту істотно не змінюється, оскільки периостальний ріст кістки в діафізі в дорослих жаб практично припиняється.
Таким чином, у ростучих кістках скелету амфібій окістя представлене складною структурою, що складається з остеогенних клітин, що знаходяться на різних стадіях диференціювання. У кістках сформованого і ріст кістяка, що закінчився, вона перетворюється в одношарову структуру, що складається з зовні однотипових фібробластоподобних клітин.
Розділ 3. Пролиферативні властивості клітин окістя в амфібій
У спеціальній літературі є нечисленні загальні відомості про розвиток кістяка довгих трубчастих кісток в амфібій (Кащенко, 1896; Eggelіng, 1911; Lubosch, 1928; Румянцев, 1958; Житников, 1980; Домашевская, 1984, 1989). При цьому фактично відсутня інформація про проліферативних потенції кліток окістя, що відіграє велику роль в апозиційному рості кістки. Для більш повного з'ясування особливостей росту і розвитку кістяка в амфібій представляється важливим вивчення не тільки структури окістя, але і проліферативних потенцій її клітин.
Дослідження проводили на личинках, маса яких складала 4-5 м, і на однорічних часночницах (Pelobates fuscus). 3 Н-тимидин уводили внутрішньочеревно однократно в дозі 74 кбк/м живої маси з наступною фіксацією періосту з підлягаючим шаром компакт стегнової кістки в 10 %-м нейтральному формаліні через 1; 24 і 48 год. (личинки) і через 1; 24 і 72 год. (жабенята).
На гістоавтографах з 3 Н-тимидином визначали (у %) відношення мічених кліток до загального числа кліток на умовну одиницю площі гістологічного зрізу, а також інтенсивність синтезу дезоксирибонуклеїнової кислоти по концентрації мітки над ядром клітки (інтенсивність мічення). Мітотичний індекс ядер обчислювався на 1000 клітин. Отримані дані обробляли методом варіаційної статистики (Терентьєв, Ростова, 1977).
У період розвитку личинки (стадія 46, 47, 49 по Н. В. Дабагяну) окістя довгих трубчастих кіст у зонах закладки кісти діафіз, метафіз і "перехідна зона" різна по товщині (кількості клітинних рядів) і функціональним проявам кліток. Так, у личинок (на стадії 46) окістя починає диференціюватися в діафізарній зоні майбутньої кісти в період, коли хрящова закладка підготовлена до заміщення. У найглибшому шарі перехондра ще до появи тут кісткової манжетки клітки здобувають характерний вид остеобластів і починають продукувати фермент - лужну фосфатазу. У цей час у земноводних відзначається типове для амфібій грибоподібне розростання хрящового епіфіза, краю якого далеко заходять на метафізи. Клітки окістя в "перехідній зоні" (на границі перихондр -і окістя) розташовані в 2-3 ряду, з яких внутрішній шар складається з остеобластів, а поверхневий - хондробластів. У кінцевому верхівковому відділі щілини, куди край кісткової трубки не доходить, утримуються тільки хондробласти (малюнок, а).
Уже через годину після введення 3 Н-тимидина індекс мічення клітин, як по шарах перихондра, так і по зонах хрящової закладки кісти, неоднаковий (малюнок, б): він, по-перше, значно вище в зоні діафізу закладки кістки і, по-друге, великий відсоток мічених ядер відзначається в його поверхневому шарі. Варто звернути увагу на те, що і показник мітотичного розподілу цих клітин вищий.
Очевидно, сумарний показник концентрації всіх клітин окістя, як і фракції в кожній зоні закладки кістки, безпосередньо зв'язаний з інтенсивністю протекаючого тут перихондрально-періостального остеопластичного процесу. Тому що в метафізарній зоні приріст йде повільно, те тут і зосереджено на еквівалентній площі мінімальна кількість проліферуючих клітин.
Приблизно така ж картина розподілу кліток у зоні метафиза зберігається і через 24 год. після введення 3 Н-тимидина (личинки досягають 47-й стадії розвитку), однак до цього часу вагомо (майже в 2 рази) зменшується інтенсивність мічення кліток поверхневого шару - свідчення розподілу, що пройшли за цей час їх, (від 13 до 27 %).
У цей час перихондральна кісткова трубка в області діафаза здобуває трохи великі розміри по довжині і масі. Якщо на попередній стадії зачаток кісткового діафізу виглядав на гістологічному зрізі як тонка (10-12 мкм) пластинка, то на стадії 47 товщина її досягає 24 мкм, і в ній уже є одиничні остеоцити.
Паралельно з цим слід зазначити кількісне збільшення мічених остеобластів (з 17 до 38 %) і незначна кількість мічених фібробластоподобних клітин. Однак якщо збільшення мічених остеобластів відбулося за рахунок дозрівання як позначених преостеобластів, так і розподілу самих остеобластів, ту утримуючу мітку фібробластоподібні клітки розподіляються в межах своєї ж популяції поверхневого шару.
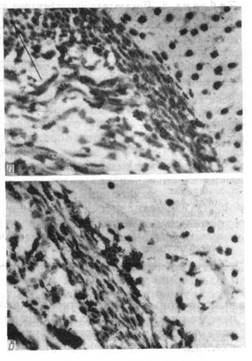
Гістоавтографи окістя стегнової кісти личинки P. fuscus на стадії 47 (по Н. В. Дабагяну). Видні численні мічені клітки окістя (а) і окістя зони діафізу (в) через 1 год. після внутріочеревного введення 3 Н-тимідину (Гемат.-еозин Делафильда; про. 40. ок. 7).
Через 48 год. після введення 3 Н-тимидина можна відзначити наступне: 1) помітно прогресує фракція мічених кліток у глибокому шарі (зона діафізу) при одночасному значимому зниженні індексу мічення кліток поверхневого шару, тобто поповнення остеобластів, очевидно, відбувається за рахунок кліток поверхневого шару періоду; 2) частина мічених кліток метафіза розділилася повторно. Мітка над ядрами деяких кліток стала на рівні тла, і вони не враховувалися при підрахунку.
Отже, сам собою напрошується висновок про те, що інтенсивність остеопластичного процесу в зростаючій закладці кістки в даний період онтогенезу тісно зв'язана з кількісною концентрацією остеогенних клітин і їх проліферуючих одиниць у кожній конкретній зоні періхондра - періосту, що в кінцевому рахунку і позначиться на росту кістякових ланок у цілому.
Для того щоб упевнитися в можливості цього, ми поставили додатковий досвід на однорічних чесночницях при тих же режимах введення 3 Н-тимидина в організмі. Перед початком викладу результатів даного дослідження відзначимо, що в річної жаби діафізарна область стегнової кістки вже цілком сформовані, ростові процеси зберігаються ще в метафізах і значно активно протікають у перехідній зоні.
На відміну від окістя більш ранніх стадій розвитку личинок у жаб на цій стадії онтогенезу остеобласті в періості діафіза вже здобувають форму подовжених кліток із щільним штриховидним ядром. Такий метаморфоз остеогенних клітин, очевидно, зв'язаний з переходом окістя діафізарної зони кістки в стан більш зрілої тихорєцької диференціювання, для якої подовжена форма остеогенних клітин є звичайною. Незважаючи на зміну форми, їхня кількість на рівній довжині періосту зберігається в межах 17-20 одиниць на 250 мкм.
Природно, виникає питання: яким шляхом поповнюється склад остеогенних клітин в окісті, за рахунок яких джерел?
При аналізі отриманих гістоавтографів виявилося, що через годину після введення 3Н-тимидина в окісті діафізу відзначаються усього одиничні (до 6%) мічені клітини серед преостеобластів. В окісті зони метафізів реєструються мічені клітини усіх видів зрілості (остеобласти, преостеобласти і фібробластоподібні клітини), однак їхній відсоток невисокий (від 5 до 7 %). У той же час у "перехідній зоні" індекси мічення ядер над цими видами кліток трохи вище, а над преостеобластами і фібробластоподобними клітками індекси мічення ядер майже в 3 рази вище, ніж в окісті зони метафіза кістки.
Через 24 год. після початку досліду в зоні діафіза як і раніше реєструються невисокі відсотки мічених ядер над клітинами - 2-4 %, у той час як над ядрами клітин метафіза і перехідної зони - значно вище. Так, над прехондробластами індекс мічення ядер - 18 %, а над преостеобластами - 53%. При цьому відзначається зниження на половину інтенсивності мітки (з 60 до 22 зерен відновленого срібла), через 24 год. ми врахували фактично популяцію поділившихся раніше позначених клітин.
У метаепіфізарних зонах під грибоподібною муфтою епіфізарного хряща продовжує існувати періостально-перихондральна ділянка, у якому на границі з гіаліновим хрящем клітки такого тихорєцького утворення диференціюються в хондробласти, а на границі з кістою і кровоносними судинами - в остеобласти. Поверхневий шар окістя, як і на попередніх стадіях, складається з волокнистого каркаса з включеними клітками різного рівня диференціювання - від фібробластоподобних до преостеобластів. Поповнення кліток у ній відбувається переважно за рахунок менш диференційованих станів, і частково за рахунок функціонально зрілих остеобластів. Тобто , окістя в земноводних тут представлена єдиним шаром: входять до складу клітини різних стадій диференціювання.
При аналізі гистоавтографов через 72 год. у зоні діафізу кістки в окісті відзначається як і раніше невисокий індекс мічення ядер клітин. Серед ядер клітин окістя зони метафізів індекси мічення ядер також невисокі.
При цьому над фібробластоподобними клітинами відзначається і розведення мітки, тобто має місце, очевидно, не тільки їхній розподіл, але і трансформація в інші ряди клітин, наприклад, за їхній рахунок йде поповнення популяції преостеобластів. Зниження індексів мічення ядер спостерігається і над ядрами клітин перехідної зони, з одного боку, за рахунок зменшення прехондробластів ми маємо збільшення відсотка хондробластів, а з іншого боку - за рахунок раніше мічених преостеобластів - значний ще відсоток мічених остеобластів (19 %).