Курсовая работа: Розвиток нервової тканини у безхвостих
А. Культура нейробластів у неорієнтованому середовищі. Нервові волокна ростуть у всіх напрямках. Б. Середовище орієнтоване шляхом розтягування, після чого нервові волокна ростуть паралельно один одному. В. Дві культури орієнтують середовище, що знаходиться між ними, і нервові волокна ростуть паралельно один одному.
Концепція "контактної спрямованості" зустрічає менше заперечень, чим дві інші гіпотези про причини росту нервових волокон у напрямку органів. Однак вона, очевидно, не дає повного пояснення спрямованого росту нервових волокон. Орієнтований субстрат може забезпечити загальний напрямок росту волокон, але навряд чи цим можна пояснити численні випадки дуже точної орієнтації волокон і встановлення специфічних функціональних зв'язків. Цілком можливо, що очна стеблинка служить механічним субстратом, що забезпечує вростання нервів у головний мозок, а субклітинної орієнтацією субстрату цілком можна пояснити ріст нервів у напрямку до кінцівки, однак подальший ріст цих нервів у мозку і кінцівці припускає існування інших, хоча, бути може, і подібних, механізмів. Мігруючі пігментні клітини, що беруть початок від нервового гребеня, можуть переміщатися уздовж ліній орієнтації субстрату, але цим не можна пояснити виникнення упорядкованих візерунків пігментації. У культурі ці клітки розташовуються на визначених відстанях друг від інша імовірно, таке їхнє розташування в культурі, а також міграція від нервового гребеня по бічних сторонах тіла тварини порозуміваються взаємним відштовхуванням кліток. Таким чином, навіть незважаючи на те, що "контактна спрямованість", можливо, визначає основні шляхи переміщення кліток до місця їхнього призначення, концентрація продуктів обміну речовин у цих клітках може служити стимулом для їхньої міграції і забезпечувати остаточний розподіл і положення кліток [6].
Заслуговують також згадування дані про існування іншого роду специфічності. Звичайно, як тільки нерви досягають кінцівки, яка розвивається, вони розподіляються в ній нормальним образом. Примітно, що якщо нерви доростають до добре розвитий передньої кінцівки, цілком позбавленої нервів, то в ній виникає той же тип іннервації, що й у молодій кінцівці, що розвивається; отже, нерви, потрапивши в кінцівку, що знаходиться на ранній або пізній стадії розвитку, розгалужуються однаково. Тенденцію нервових волокон рости уздовж вже існуючих кровоносних судин можна пояснити тим, що це шлях найменшого опору. Остаточно характер іннервації кінцівки, очевидно, встановлюється в результаті взаємодії рано розвиваються м'язів з рано розвиваються нервовими волокнами. Ця можливість найкраще виявляється при розгляді питання про вплив кінцівки на нервову систему і про специфічність нейронів.
5. Дія периферичних областей на нерви
Дуже сильний вплив на нервову систему, що розвивається, роблять структури, з якими в неї установлюється функціональний зв'язок. Це знаходить своє відображення в розмірах нервів і гангліїв. Нерви кінцівок, що містять як рухові, так і чуттєві волокна, крупніше інших спинальних нервів; ганглії нервів кінцівок крупніше, ніж інші спинальні ганглії. Видаляючи кінцівку на одній стороні зародка і залишаючи кінцівку протилежної сторони недоторканої, можна показати, що велика величина нервів і гангліїв безпосередньо зв'язана з периферичним функціональним навантаженням на нерви.
На ранніх стадіях розвитку вентролатеральна частина спинного мозку диференціюється в рухову область, і волокна з нейробластів цієї області іннервують м'яза тіла і кінцівок. Спинальні ганглії містять клітинні тіла чуттєвих нервів, і рефлекторна дуга замикається через спинний мозок. Очевидно, що між кількістю інервованої тканини і диференціацією клітин у нервовій системі існує прямий зв'язок. Цей висновок підтверджується результатами досліду з пересадженням кінцівки в нове положення. У такому досліді пересаджена кінцівка інервується новими нервовими відростками. У результаті кількість волокон у цих нервах збільшується і наближається до нормального для волокон рухового нерва [5].
У більшості хребетних кінцівка індукує утворення різного числа рухових нейронів у спинному мозку і чуттєвих нейронах у спинальних гангліях. Установлено кількісні співвідношення між периферичним навантаженням і диференціюванням нейронів в амфібії.
Кінцівка, що розвивається, створює комплексну дію на спинний мозок і ганглії. Присутність кінцівки, очевидно, викликає посилену проліферацію кліток, а також збереження кліток, що звичайно дегенерують. Відомо, що загибель клітин і нормальне явище в розвитку різних систем, у тому числі і центральної нервової системи. Присутність власної або пересадженої кінцівки запобігає звичайній дегенерації деяких нейронів у центральній нервовій системі. Відсутність кінцівки або зменшення периферичної області кінцівки приводить до збільшення кількості клітин, що дегенерують. Ці фактори при незалежній або спільній дії можуть забезпечити відповідність величини нервових, центрів їхнім периферичним зв'язкам.
Вплив периферичних областей на диференціювання клітин нервових центрах, цілком ймовірно, здійснюється через нервове волокно. При видаленні кінцівки, що розвивається, від задньої кінцівки від нейробластів у спинному мозку, як звичайно, відходять цитоплазматичні волокна. Вони ростуть у напрямку до культи і розташовуються в тканині рубця безладним чином. Протягом декількох днів ці волокна, що не прикріпилися, атрофуються, а їхні клітинні тіла у вентральній частині спинного мозку дегенерують. Очевидно, для нормального розвитку нейрона необхідно, щоб аксон нерва, що розвивається, установив контакт із визначеною структурою. Областю контакту, видимо, є конус росту волокна; якщо контакт не установився, те відповідна інформація якимсь образом передається тілу клітки, центрам її метаболічної і синтетичної активності, у результаті чого припиняється диференціювання або відбувається дегенерація. Отже, установлення периферичних зв'язків досить істотно для підтримки нервових кліток, що розвиваються, у визначеному стані. Збереження і виживання нервових кліток, можливо, залежить також і від їхніх взаємин один з одним. Процес дегенерації може переходити з одного нейрона через аксон і синапс на інший нейрон, що спричиняє дегенерацію цього другого нейрона; таке явище називають міжнейронною дегенерацією; його, зокрема, можна спостерігати в слуховій і зорової системах. Наприклад, навіть саме незначне ушкодження сітківки приводить до помітної атрофії зорових часток середнього мозку шляхом міжнейронної дегенерації.
На диференціюванні клітин у нервових центрах, крім зовнішніх факторів, що діють через нервові волокна, впливають також гуморальні фактори. Дослідження іn vіtro показали, що фактор росту скоріше діє на тіло клітини, а не на аксон. Дією саме на тіло клітки порозумівається, очевидно, посилене розвиток нервових клітин під впливом тиреоїдного гормону. У процесі нормального розвитку гуморальні агенти (гормони) по кровоносних судинах можуть попадати в різні частини нервової системи і безпосередньо впливати на що розвиваються нейробласти [4].
Зв'язок нервових центрів, що розвиваються, з периферичними областями залишається багато в чому нез`ясованим. Та обставина, що зовнішні фактори впливають на диференціювання нейронів, яким би не був механізм цього впливу, можливо, ще раз підкреслює важливу роль, що відіграють різні форми індукційних взаємодій у процесі прогресивного диференціювання систем, що розвиваються.
6. Розвиток функціональних зв'язків
Спинний мозок зародка характеризується лабільною організацією. В амфібій він організований таким чином, що 3-5-і сегменти іннервують передні кінцівки, а 6-8-і сегменти - м'яза, розташовані за цими кінцівками. Ці дві ділянки на ранніх стадіях розвитку можна поміняти місцями, і вони замінять один одного. Як показано на рис. 10, у зародка можна видалити3-5-і сегменти і замінити їхній 6-8-ми сегментами, узятими в іншого зародка. При цьому функція передньої кінцівки цілком зберігається, а це означає, що на ранніх стадіях розвитку 6- 8-і сегменти лабільні і їх можна змусити функціонувати по типу3-5-х сегментів.
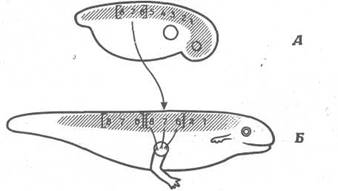
Рис. 10. Заміна 6-, 7- і 8-го сегментів спинного мозку 3-, 4- і 5-м сегментами. Зачаток передньої кінцівки залишається в колишнім положенні. Кінцівка, що розвивається, притягає нерпи з пересаджених 6-, 7- і 8-го сегментів і нормально функціонує. Отже, нове середовище викликало розвиток 6-, 7- і 8-го сегментів по типу 3-, 4- і 5-го сегментів. А. Пересадження 6- 7- і 8-го сегментів на місце 3-, 4- і 5-го. Б. Пересаджені сегменти нормально функціонують.
Принцип лабільної організації прикладемо також до 3-5-му нервів. Додаткова брунька кінцівки, трансплантована поруч з нормальної, часто інервується відгалуженням 5-го нерва. У цьому випадку додаткова кінцівка функціонує нормально; якщо ж вона інервується 6-8-ми нервами без відгалуження від 5-го нерва, то функціонувати нормально вона не може. У той же час однієї гілочки кожного з 3-5-го нервів досить для нормального функціонування передньої кінцівки. Таким чином, на ранніх стадіях розвитку всі розходження в організації цих трьох нервів носять лабільний характер. Нагадаємо, що спінальні нерви утворять сплетення в основі кінцівки, так що сегменти, що іннервують кінцівку, мають тенденцію функціонувати як єдина група [5].
Лабільність нервів виявляється в явищі гомологічної функції. Якщо м'яз кінцівки личинки амфібій, пересаджений поруч з нормальною кінцівкою, інервується відгалуженням від нерва кінцівки (наприклад, відгалуженням від 5-го спінального нерва), то пересаджений м'яз завжди буде скорочуватися одночасно з таким же м'язом у нормальній кінцівці. Якщо пересадити в такий же спосіб цілу кінцівка, то кожен м'яз останньої буде скорочуватися одночасно з її гомологом у нормальній кінцівці і руху додаткової і нормальної кінцівок будуть синхронними. Ще більш дивно, що якщо додаткову кінцівку пересадити так, що її полярність стає протилежної полярності нормальної кінцівки, розташованої на тій же стороні, те їхнього руху будуть дзеркально відбивати один одного. Отже, нерви, що постачають гомологічні м'яза, тобто м'яза, що володіють тотожною конституціональною специфічністю, стають функціонально зв'язаними, хоча такі функціональні ефекти можуть виявитися марними або навіть шкідливими для тварини. Описане явище гомології функцій показує, що м'яз реагує на особливий вид стимулу незалежно від специфічності її нерва, що іннервує, а це означає, що м'яз, або периферична область, відіграє важливу роль у встановленні специфічних функціональних зв'язків у нервовій системі.
6. Специфічність відносин центра і периферії
Численні дані показують, що в ході диференціювання нейрони набувають визначеної біохімічної специфічності, що складає основу складних функціональних взаємозв'язків між нервовими центрами і периферичними областями. Ця проблема була найбільше повно вивчена на прикладі зорової системи хвостатих амфібій. Як у дорослих, так і в зародків цих тварин можна вирізувати ока, повернути їх і знову посадити на місце; при цьому зоровий нерв відновлюється. Однак одержувані результати різко розрізняються в залежності від того, на якій стадії була проведена операція. Так, до стадії пізньої хвостової бруньки око можна повернути на 180° без яких-небудь серйозних порушень зору або очноорухливої координації. Пізніше, однак, така ж операція різко порушує очних реакцій тварини [1].
Справедливість представлення про специфічність зв'язків між чуттєвим центром і периферичною областю підтверджується також даними, отриманими для систем, що відрізняються від зорової, котра, являє собою центральну систему, оскільки сітківка утвориться з частини нервової пластинки. Існування специфічного зв'язку між нервом і периферичною областю було також показане в досвідах зі шкірними нервами. Якщо в пуголовка жаби вирізувати дорзовентральну смужку шкіри, повернути неї на 180° і знову посадити на місце, вона приживляється і буде самодиференціюватися, так що після метаморфоза на спині тварини буде шматок світлої шкіри черева, а на череві - шматок темної шкіри спини. Якщо подразнювати ділянку темної шкіри на животі, то тварина намагається змахнути подразник зі спини або усунути подразник з живота, якщо подразнювати ділянку світлої шкіри на спині.
Дані, отримані в таких дослідах, пояснити не дуже легко, і будь-яке їхнє пояснення в даний момент буде чисто умоглядним. Однак пропоноване нижче пояснення співпадає з відомими фактами і показує, які процеси можуть брати участь у встановленні численних і різноманітних зв'язків у нервовій системі (рис. 11). Цілком імовірно, що тип центральних синаптичних зв'язків, утворених нервовими волокнами, визначається, принаймні частково, властивостями периферичної області, з яким зв'язані ці волокна. Таким чином, можна представити, що, як тільки конус росту волокна, що розвивається, досягає шкіри, в області контакту між нервом і шкірою відбувається індукційна взаємодія, за допомогою якого шкіра додає нервові особливу біохімічну специфічність, що визначає виникнення центральних зв'язків шляхом більш тонких індукційних впливів на рівні нейронів. Якщо розглянути з цих позицій зорову систему, то виявиться, що на визначеній стадії розвитку сітківка стає поляризованої і на більш пізніх стадіях зв'язок між будь-якою частиною сітківки і зоровою часткою закріплюється. Таким чином, можливо, що в процесі розвитку сітківка диференціюється до такого стану, коли кожна сама маленька ділянка її володіє особливою біохімічною специфічністю. Досягши зорової частки, волокна специфічних кліток наділяють відповідною специфічністю центральні нейрони. Отже, аксони, що проникають у відповідну частину зорової частки, можуть сприяти виникненню специфічності в тканині центральної нервової системи, а після цього зв'язок між сітківкою і зоровою часткою жорстко фіксується, подібно зв'язку між шкірою і нервом. Тут важливо нагадати, що порушення очнорухливих реакцій після повороту ока, що свідчить про втрату лабільності системи "сітківка - зорова частка", виникає в той час, коли перші волокна зорового нерва досягають зорової частки. У процесі розвитку відбувається просторове диференціювання нервової системи.
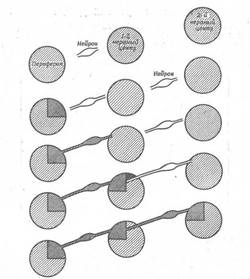
Рис. 11. Схема послідовних процесів, що приводять до виникнення специфічності нейронів і нервових центрів.
Специфічність, очевидно, уперше виникає в периферичній області. Потім зростаючі нейрони, установлюючи функціональні зв'язки з периферичною областю, також здобувають відповідну специфічність. Після цього вони передають свою специфічність нервовим центрам. Таким шляхом різні нейрони і нервові центри здобувають у ході розвитку взаємозалежну анатомічну і функціональну специфічність.
Нейрони з різних частин головного мозку часто розрізняються за формою і розміром. У мозку відбувається диференціальний розподіл холинэстеразы і катехоламинов, а тканина центральної нервової системи істотно відрізняється від периферичної по антигенному складі [8].
Становлення особливої біохімічної специфічності зв'язків між центральною і периферичною областями не означає, що внутрішня організація частин нервової системи цілком залежить від тонких індукційних впливів, що торкаються специфічність нейронів; воно не виключає необхідності направляти ріст волокон і не є незалежним від таких факторів, як тимчасова послідовність розвитку і відповідні просторові зв'язки.
Висновок
Підводячи підсумки, можна сказати, що загальні принципи розвитку організму на ранніх стадіях застосовні також до розвитку нервової системи.
Як для центральної нервової системи, так і для периферичних нервів характерні вихідна лабільність і взаємодія, що детермінує, з периферичними областями.
Розвиток специфічної функції, що починається з найпростішої форми організації і перехідне до самої витонченої взаємодії між нейронами в соматосенсорної області кори головного мозку, характерно для нервової системи. Відповідно м'язові рухи і пристосування тварини до середовища розвиваються від загальних до більш специфічного. Індукційні й інші взаємодії кліток і вироблення специфічних біохімічних властивостей у нервовій системі й у зв'язаних з нею структурах уражають своєю складністю і для їхнього остаточного аналізу буде потрібно застосування безлічі різноманітних методів.
Список літератури
1. Александровская О.В., Радостна Т.П., Козлов НА. Цитология, гистология и эмбриология. - М.: ВО «Агропромиздат», 1987 - 448 с.
2. Белоусов Л. В. Введение в общую эмбриологию: Изд-во Московского университета, 1980. — 210 с.
3. Бодемер Ч. Современная эмбриология. — М.: «Мир», 1971. — 446 с.