Курсовая работа: Сенсорные процессы и восприятие у животных
Введение
Способность животных реагировать на изменения среды зависит от сенсорных процессов, предназначенных для обнаружения этих изменений, начиная от простого обнаружения разницы температуры или силы света и кончая распознаванием сложных паттернов. В настоящей главе мы рассматриваем основные сенсорные модальности у животных и исследуем природу восприятия.
1. Хеморецепция и терморецепция
Хеморецепцией называется способность распознавать химические вещества и определять их концентрацию. Ею обладают даже очень примитивные формы живого. Собственно говоря, любую нервную клетку можно считать хеморецептором, так как она специфически реагирует на вещества, выделяемые другими нервными клетками. Механизмы хеморецепции включают распознавание специфических молекул рецепторными участками на клеточных мембранах. Не совсем ясно, основано ли это на химическом действии, на форме молекулы или на том и другом одновременно. Например, мы не знаем, какая общая черта сахара и сахарина делает оба этих вещества сладкими для падальных мух, крыс, обезьян и человека.
Хеморецепторы делятся на экстероцепторы и интероцепторы. Первые реагируют на присутствие химических веществ во внешней среде, а вторые - на соединения, циркулирующие в жидкостях тела, в частности на окись углерода, питательные вещества и гормоны. И вкус, и обоняние зависят от хеморецепторов. В традиционном понимании обоняние различает низкие концентрации веществ, содержащихся в воздухе, а вкус возникает от прямого соприкосновения с относительно высокими концентрациями химических веществ. Однако в обоих случаях на рецептор действуют растворенные вещества; у некоторых животных, например у водных обитателей, разделить эти чувства трудно. Тем не менее у многих животных имеется неврологическое различие, которое состоит в том, что одни нервы связаны с обонянием, т. е. обнаружением низких концентраций, а другие проводят вкусовые сигналы от разных рецепторов, специфически реагирующих на относительно концентрированные химические вещества. Так, у падальной мухи хеморецепторы антенн распознают малые количества веществ в воздухе, а хеморецепторы на лапках способны обнаруживать соль, сахар и чистую воду. У позвоночных вкусовые сигналы передаются по лицевому и языкоглоточному черепномозговым нервам, а обонятельные - по обонятельному нерву.
Одна из наиболее исследованных обонятельных систем у беспозвоночных связана с восприятием феромона ухаживания у тутового шелкопряда. Феромон - это соединение или смесь соединений, выделяемая организмом в окружающую среду и вызывающая специфическую поведенческую или физиологическую реакцию у воспринимающего организма того же вида. Таким образом, феромоны - это химические посредники, участвующие, вероятно, в самой примитивной форме общения животных. Первым химически идентифицированным половым феромоном был феромон тутового шелкопряда; это вещество получило название «бомбикола». Его секретируег
абдоминальная железа самок, которые таким способом могут привлекать самцов на расстоянии нескольких километров. Химическое строение бомбикола известно, он был синтезирован, испытывался на самцах этого вида или применялся для стимуляции изолированных антенн, к рецепторам которых были подведены электроды. Полную физиологическую реакцию вызывает только бомбикол, а более слабый ответ возникает и на некоторые очень близкие к нему вещества. Это говорит о большой специфичности. Еще примечательнее то, что полную реакцию может вызвать всего одна молекула бомбикола. Поведенческий ответ самца шелкопряда состоит в том, что он летит против ветра, ориентируясь по молекулам бомбикола и, найдя самку, спаривается с ней. Такой тип ориентации на химический стимул очень распространен у насекомых.
Насекомые обладают разными типами обонятельных сенсилл. На их поверхности обычно находятся многочисленные мелкие поры, которые заканчиваются трубочками, наполненными жидкостью. Дендриты рецепторных клеток заходят в сенсиллы, а их аксоны идут прямо в мозг. Взвешенные в воздухе молекулы феромона попадают в поры сенсиллы и через них проникают в заполненную жидкостью внутреннюю часть, где приходят в соприкосновение с рецепторной мембраной.
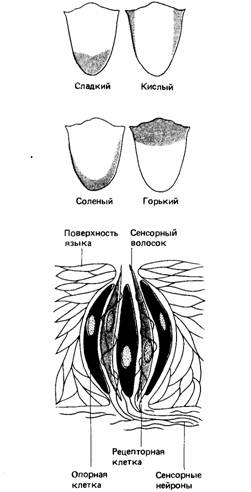
У позвоночных обонятельные рецепторы представляют собой сенсорные нейроны первого порядка, дендриты которых в виде ресничек погружены в слизистый слой. Аксоны этих нейронов идут к обонятельной луковице, где образуют синапсы с нейронами второго порядка, аксоны которых образуют обонятельный тракт, входящий в передний мозг. Вкусовые рецепторы состоят из чувствительных клеток, которые обычно расположены группами, называемыми сосочками. Чувствительные клетки плотно прилегают к сенсорным нервным волокнам, которые остаются неизменными, тогда как вкусовые клетки каждые несколько дней замещаются новыми.
У млекопитающих четыре основных вкусовых ощущения - кислого, горького, соленого и сладкого.

Рис. 1. Схематическое изображение обонятельной сенсиллы бабочки.
У людей к каждому из них чувствительны разные части языка. Кислое воспринимается боковыми сторонами ближе к корню, горькое - самой задней областью, соленое - боковыми сторонами ближе к переднему краю, а сладкое -кончиком языка. Вкусовые качества пищи зависят одновременно от ее вкуса и запаха. Это легко показать, попросив испытуемого определить с закрытыми глазами разницу между мелкими кусочками яблока и лука, положенными ему на язык. Большинство людей легко их различают, если имеют возможность дышать через нос, но не способны сделать это с зажатым носом.
Сделано много попыток выделить основные запахи по примеру четырех основных вкусовых ощущений. Наиболее распространенная классификация, приводимая в табл. 1, принадлежит Дж. Эймуру. Еще будучи студентом Оксфордского университета Эймур заметил большие различия в химическом строении одинаково пахнущих веществ.
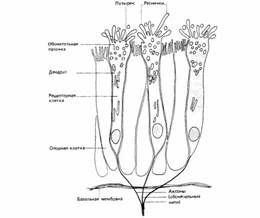
Рис. 2. Схематическое изображение обонятельной сенсорной. Три сенсорные клетки окружены опорными клетками
Он исследовал более 600 органических соединений с детально описанными запахами и в 2 г. опубликовал свою стереохимическую теорию обоняния, в которой все они охарактеризованы на основе семи первичных запахов, перечисленных в табл. 1. Теория Эймура классифицирует химические вещества по форме и величине их молекул. Он постулирует существование семи основных типов рецепторов, характеризующихся особыми рецепторными участками, в которые могут входить молекулы определенной формы. Так, молекулы веществ с камфарным запахом приблизительно сферические, а у веществ с мускусным запахом они имеют форму дисков. Теория Эймура получила некоторое подтверждение в дальнейших исследованиях, но все же остается противоречивой.
К хеморецепции несколько близка терморецепция, которая, вероятно, имеется у большинства животных, но изучена у относительно немногих. Чувствительные к температуре нервные окончания обнаружены у разных насекомых. Например, у таракана Periplanetaтерморецепторы на лапках воспринимают температуру почвы, а на антеннах - температуру воздуха. У рыб терморецепторы находятся в коже, боковой линии и головном мозгу, что делает их очень чувствительными к изменениям температуры. Показано, что сомы реагируют на перепады температуры менее 0.1C. Многие пресмыкающиеся обладают хорошо развитой температурной чувствительностью и терморецепторами в головном мозгу и коже. У ямкоголовых змей на голове имеются так называемые лицевые ямки, чувствительные к инфракрасному излучению и устроенные так, что животное способно к дирекциональной температурной чувствительности тела.
Покровы птиц, как полагают, бедны терморецепторами, за исключением языка и клюва некоторых видов. У голубей терморецепторы головного мозга влияют на поведение и расположение оперения, а другие - в спинном мозгу - управляют дрожью и одышкой. У млекопитающих специальные тепловые и холодовые рецепторы распределены по коже, причем тепловые обычно залегают глубже Холодовых. Имеются также рецепторы в глубине тела, которые могут вызывать дрожь, даже если температура кожных и мозговых рецепторов не изменяется. Терморецепторы спинного мозга влияют на дрожь, одышку и кровоток, причем эти же функции дублируются терморецепторами гипоталамуса. У млекопитающих в целом наблюдаются самые развитые формы терморегуляции. Их головной мозг получает сигналы от многих частей тела: их интеграция ведет к соответствующему включению различных согревающих и охлаждающих механизмов.
Табл. 1. Первичные запахи с химическими и бытовыми примерами
Рис. 3. Распределение и строение вкусовых почек человека. | Первичный запах | Химический пример | Бытовой пример |
| Камфорный | Камфора | Репеллент против моли | |
| Мускусный | Пентадеканолактон | Масло из дудникова корня | |
| Цветочный | Фенилэтилметилэтилкарбинол | Розы | |
| Мятный | Ментон | Мятные леденцы | |
| Эфирный | Этилендихлорид | Пятновыводитель | |
| Едкий | Муравьиная кислота | Уксус | |
| Гнилостный | Бутилмеркаптан | Тухлое яйцо |
2. Механорецепторы и слух
Звук возникает вследствие ничтожных изменений давления, вызываемых источником колебаний в воздухе или в воде. Звуковые рецепторы в принципе представляют собой механорецепторы с быстрым восстановлением чувствительности, что делает их восприимчивыми к колебаниям.
У множества членистоногих описаны чувствительные к вибрации волоски и рецепторы в суставах конечностей. У падальных мух в некоторых суставах антенн рецепторы, называемые джонстоновыми органами, могут реагировать на колебания частотой до 500 Гц. У комаров орган такого же типа сигнализирует о направлении звука. Гладыш Notonectaлокализует добычу по волнам, распространяющимся на поверхности воды. Рецепторы, расположенные на его лапках, максимально чувствительны к колебаниям частотой - Гц. Пауки отличают живую добычу от мертвой по вибрации паутины. Некоторые позвоночные обладают механорецепторами, способными воспринимать колебания субстрата. Такие рецепторы найдены в коже змей и в суставах ног у кошек и уток.
Слуховые системы животных, несмотря на разнообразие, имеют некоторые общие черты. Так, например, их периферическая часть преобразует звуковое давление в колебательное движение. Рецепторы превращают это движение в нервные импульсы, которые декодируются ЦНС. Один из самых простых типов периферических устройств известен у бабочек совок. У них имеются два «уха», каждое из которых состоит просто из тимпанальной мембраны по бокам груди и двух рецепторных клеток, погруженных в полоску соединительной ткани. Это поразительно простое «ухо» позволяет бабочкам слышать ультразвуковые сигналы охотящихся летучих мышей. В серии изящных опытов Редер показал, как это происходит.
Один рецептор, называемый клеткой A1 , чувствителен к слабым звукам и реагирует на сигналы летучих мышей на расстоянии около 30 м, т. е. слишком далеким для того, чтобы мышь обнаружила бабочку. Частота импульсов, идущих от клетки A1 , пропорциональна силе звука, и поэтому бабочка может определить, приближается ли летучая мышь. Сравнивая время прихода и интенсивность стимула в обоих органах слуха, бабочка узнает о направлении полета мыши. Разница возникает потому, что тело бабочки заслоняет звук от одного «уха» больше, чем от другого. Может быть также определена относительная высота полета летучей мыши: когда она выше насекомого, звук, приходящий к тимпанальным органам, ритмически прерывается взмахами крыльев бабочки; когда мышь находится ниже бабочки, этого не происходит.

Рис. 4. Тимпанальный орган совки. Колебания тимпанальной мембраны воспринимаются сенсорными нейронами А1 и А2 .
Клетки А1 заранее предупреждают бабочку о приближении летучей мыши и могут позволить ей улететь до того, как мышь ее обнаружит. Удаляясь по прямой, бабочка становится наименьшей мишенью, потому что ее крылья обращены к мыши краями, а не плоскостью; она добивается этого, просто разворачиваясь так, чтобы уравнять звук, достигающий обоих «ушей». Но если летучая мышь обнаруживает бабочку, та не может спастись от нее, просто улетев, так как летучая мышь движется гораздо быстрее. Поэтому бабочка уклоняется от хищника, когда он приблизится к ней на 2-3 м.
Клетка А2 генерирует нервные импульсы только в ответ на громкие звуки. Она начинает реагировать, когда летучая мышь близко, и, вероятно, ее импульсы прерывают действие механизмов ЦНС, управляющих полетом. В результате он становится беспорядочным, и бабочка устремляется к земле. Наблюдения показывают, что с помощью такого уклоняющегося движения бабочки спасаются от летучих мышей как раз в тот момент, когда они приближаются на расстояние непосредственного нападения. Таким образом, исследования Редером слуха у бабочек служат прекрасной иллюстрацией не только механизмов работы простого уха, но и того, что сенсорный аппарат животного тонко адаптирован к его экологии.
Звук обладает многими свойствами, на которые животное может реагировать. Когда он проходит через среду, ее частицы движутся взад и вперед, создавая осциллирующие волны давления. Величина этих волн определяет интенсивность воспринимаемого звука. Скорость его зависит от плотности среды распространения и не зависит от интенсивности. В воздухе звук распространяется со скоростью около 340 м в секунду, причем в горячем воздухе - быстрее, чем в холодном. В воде его скорость приблизительно в четыре раза выше, чем в воздухе.
Если мы изобразим звук в форме волн, как на рис. 6, то расстояние между последовательными пиками, называемое периодом, обратно пропорционально частоте, т. е. числу пиков в единицу времени. Частота звука измеряется в герцах, или в циклах, в секунду. Простейший звук - чистый тон, единственная частота которого субъективно воспринимается как высота. Естественные звуки редко бывают чистыми тонами и состоят главным образом из ряда смешанных частот. Когда сложный звук разлагают на составляющие частоты, получают так называемый звуковой спектр. Орган слуха может быть чувствителен к широкому диапазону частот; например, тимпанальный орган саранчи реагирует на частоты от 0 до 000 Гц. Когда слуховой рецептор воспринимает узкий частотный диапазон, говорят, что он «остро настроен».
--> ЧИТАТЬ ПОЛНОСТЬЮ <--