Научная работа: Клеточные и молекулярные механизмы депривационных изменений
В экспериментах, описанных нами до сих пор, основным принципом было то, что два глаза конкурируют друг с другом за связи и территорию в ЛКТ и в слое 4 первичной зрительной коры, исходно имея примерно равные возможности. Ракич с коллегами использовали другой подход для изучения того, как соседние группы клеток с хорошо определенными свойствами распределяют свои окончания и определяют свои мишени в процессе развития без конкуренции друг с другом.
Как мы уже писали выше, крупноклеточная (M, magnocellular) и мелкоклеточная (Р, parvocellular) системы располагаются в различных слоях ЛКТ и зрительной коры. При окраске отдельных аксонов M и Ρ клеток по мере того, как происходит их рост в ЛКТ во время развития, было показано, что аксоны извне приходят строго в M и Ρ слои, где они образуют характерные неперекрывающиеся паттерны ветвления. M волокна заканчиваются только в слоях 1 и 2 ЛКТ, а Р волокна — в слоях 3, 4, 5 и 6 (рис. 3), без перекрытия.
Таким образом, когда два глаза образуют свои связи, можно предположить, что конкуренция играет основную роль в разделении поступающей от них сходной информации об окружающем мире. На самом деле M и Р системы переносят абсолютно различные виды информации. Их соединения (подобно тем, что происходят при образовании пятен и полосок в зрительной зоне 2)образуются по другим принципам, в которых конкуренция играет далеко не ведущую роль.
Другим примером связей, которые образуются без участия конкуренции, является развитие карт ориентации в зрительной коре котенка. Стоит отметить, что результаты подобного рода укрепляют уверенность в том, что не все, что происходит в нервной системе, или даже на рынке может быть списано на простую конкуренцию.)
Критические периоды развития слуховой системы
Результаты, полученные при изменении восприятия зрительной информации у котят и незрелых детенышей обезьяны, имеют большое количество приложений для понимания функционирования нервной системы. Интересным примером является то, как происходит адаптация слуховой системы во время ее критических периодов, что показано в экспериментах Кнудсена и его коллег на амбарной сове (barnowl). Ранний слуховой опыт формирует особенности настройки нейронов четверохолмия амбарной совы в зависимости от частоты. Следующий пример показывает, как изменения в восприятии зрительной информации в ранние периоды жизни могут повлиять на представление слуховой системы в головном мозге амбарной совы.

Рис. 3. Отсутствие конкуренции за территорию между крупно- и мелкоклеточными аксонами клеток сетчатки и ЛКТ. Помеченные нервные окончания волокон зрительного нерва в слое M и Ρ ЛКТ обезьяны продолжают расти на протяжении с 95 до 135 дня эмбриональной жизни (с Е95 до Е135). С самого начала развития аксоны ограничены только своими собственными слоями и не распространяются в соседние слои ретракции. Таким образом, МиР волокна не конкурируют за пространство.
Слуховой и зрительный опыт у новорожденных амбарных сов
Сова способна поворачивать голову точно в направлении того места, откуда раздается звук (так как она должна ловить слабо пищащую мышь). Горизонтальная локация звука осуществляется при помощи измерения разницы во времени между правым и левым ухом (interauraltimedifference) — то есть по задержке между звуковыми волнами, поступающими сначала в одно ухо, а потом — в другое. (Сова также может принимать во внимание интенсивность звука в обоих ушах для оценки вертикальной позиции: асимметрично расположенные группы перьев на ее морде отражают звук, идущий сверху, в одно ухо, а снизу — в другое.)
Другой возможностью для совы определить положение и траекторию движения мыши является зрение. На рис. 4А показано, что у нормальной взрослой совы нейронные карты пространства для зрения и слуха (neuralmapsforvisualandauditoryspace) согласованы в одном слое зрительной области четверохолмия (tectum), которая соответствует зоне верхнего двухолмия у млекопитающих. Подобное картирование осуществляется при помощи измерения ответов отдельных нейронов четверохолмия на звуки, идущие от различных участков, и световые стимулы, предъявляемые в различных участках зрительного поля.
В серии экспериментов совят выращивали со смещенными на 23 градуса вправо или влево зрительными полями, для чего на глаза им помешались специальные призмы (рис. 5.А). Это сдвигало изображение зрительного поля на сетчатке и, следовательно, его проекцию в четверохолмии таким образом, что уже не было согласования между зрительной и слуховой картами (рис. 5.В, С). На протяжении следующих 6-8 недель происходило смещение слуховой пространственной карты, и она снова соответствовала зрительной карте. Пластичность в критический период, таким образом, позволяла проводить «тонкую настройку» функций коры в результате приобретаемого совой опыта и знаний об окружающем мире. В результате этого сова начала ориентировать свои глаза по направлению к источнику звука несмотря на искажающие призмы.
На более поздних этапах эксперимента призмы удаляли с глаз совы. Теперь вновь возникал дисбаланс между зрительной и слуховой картами. В том случае, если совы были моложе 200 дней в тот момент, когда удаляли призмы, слуховая карта во второй раз смещалась, возвращаясь к своему исходному положению, которое соответствует зрительной карте (рис.5., также см. рис. 6А).
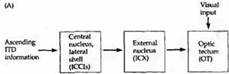
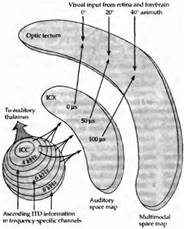

Рис. 4. Наложение друг на друга слуховой и зрительной карт пространства в области четверохолмия амбарной совы. (А) Восходящий слуховой путь, направляющийся к зрительным холмикам. Слуховые нейроны внутреннего (ICC) и наружного (ICX) ядер нижнего холмика имеют тонотопическую организацию. Они проецируют отростки в области зрительных холмиков в определенной последовательности. Слуховая карта пространства зависит от различий во времени между поступлением звуков в оба уха. Слуховые и зрительные карты пространства тесно связаны друг с другом. Таким образом, нейроны, сигналы которых регистрируются в области, отмеченной как «0 s (0 мкс)», отвечают на зрительные и слуховые стимулы, расположенные прямо перед совой. ITD — интерауральная разница во времени. (В) График ответов на интерауральную разницу во времени, полученный у молодой совы в возрасте 60 дней. Различие во времени между двумя звуками показано по оси x (слева направо). Нейроны, которые отвечают на интерауральную разницу в 0 мкс, лучше всего реагируют на стимулы, находящиеся непосредственно перед животным, и связаны с рецептивным полем, расположенным в центре зрительного поля. Звуки, идущие слева или справа, достигают ушей совы с задержкой, активируя нейроны, имеющие различные кривые ответов, пики которых отличаются от 0 мкс и которые также согласованы со зрительными стимулами.
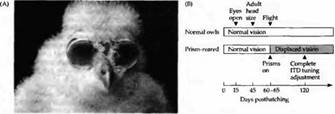
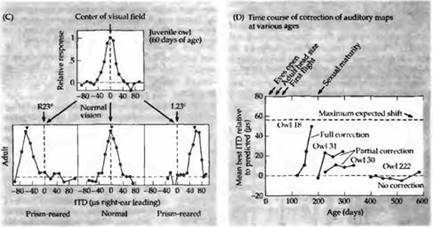
Рис. 5. Сдвиг слухового рецептивного поля после помещения на глаза сов призм во время критического периода. (А) Совята с очками, состоящими из призм, вызывающих смещение зрительного поля на 23 градуса вправо (или влево, в зависимости от очков). (В) Стадии развития, когда призмы помещались на глаза совят. (С) Аудиометрические кривые, показывающие ответы на звуки, имеющие различный интерауральный временной интервал. У нормальной молодой совы в возрасте 60 дней ответы нейронов аналогичны представленным на рис. 25.19: интерауральная разница во времени (ITD), равная 0 мкс, представляет центр зрительного поля. При выращивании животного с данными призмами происходило смещение кривых. Зрительные рецептивные поля смещались вправо или влево на 23 градуса. Теперь наибольшие ответы слуховых нейронов регистрировались с той ITD, которая соответствовала смещенным зрительным полям. При этом вновь восстанавливалось соответствие между зрительной и слуховой картами. (D) Временной ход сдвига слуховых кривых у 3 незрелых совят. У взрослой совы (сова 222) не происходило какой-либо коррекции.

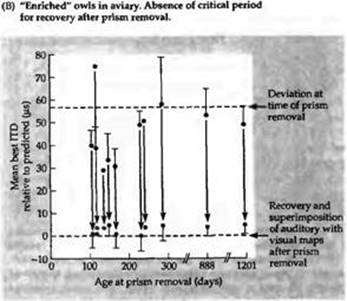
Рис. 6. Эффект обогащенного опыта в ранние периоды развития на формирование слуховой системы. (А) Результаты экспериментов с совятами, содержащимися в клетках, как показано на рис. 5. У двух совят зрительная и слуховая карта вновь пришли в соответствие (пунктирная линия), когда призмы убрали в возрасте 200 дней. Животное, у которого призмы были удалены в возрасте 219 дней (черная линия), не смогло адаптироваться к новым условиям. Карта зрительного поля вернулась к своему исходному расположению, в то время как слуховое пространство так и осталось смещенным на 23 градуса. Таким образом был определен критический период для коррекции слухового поля у сов после удаления призм. (В) Результаты экспериментов на совятах, имеющих обогащенный сенсорный опыт в раннем периоде жизни. У этих животных происходил обычный сдвиг слуховых карт и параметров ответов при помещении на их глаза призм в раннем возрасте. Восстановление после снятия призм не имело критического периода. После снятия призм происходило возвращение слуховых и зрительных карт в согласованное положение, даже если призмы снимали значительно позже 200 дней жизни.
Литература
1. Wiesel, T. N., and Hubel, D. H. 1965. / Neuro-physiol. 28: 1029-1040.
2. Guiilery, R.W., and Stelzner, D.J. 1970. J. Сотр. Neuroi. 139: 413-422.
3. Humphrey, A. L. et al. 1985. J. Сотр. Neuroi. 233: 159-189.