Реферат: Гемоглобин эритроцитарных мембран человека
DG= 2,3 RTlg(KR /KT ) кДж/моль. (7)
Для гемоглобина человека при 37°С эта "свободная энергия кооперативности" равна 5,61 кДж/моль. В физиологических условиях при отсутствии кислорода лишь ~ 3·10-5 % молекул гемоглобина находятся в R-форме, а в условиях полного насыщения кислородом лишь ~ 7-10-3 % — в Т-форме. [2]
Изменения электронной и пространственной структуры гемоглобина в процессе оксигенации На рис.4 схематически показаны электронная структура железа гема, положение атома железа относительно плоскости порфиринового кольца гема, спектральные и магнитные характеристики молекул в различных состояниях молекулы гемоглобина: дезоксигемоглобин, оксигемоглобин и ферригемоглобин. Следует подчеркнуть, что во всех случаях речь идет о равновесных состояниях молекул белка. Мы увидим далее, что переход из одного состояния в другое требует значительного (в молекулярных масштабах) времени, в течение которого система проходит через несколько неравновесных состояний, заметно отличающихся по своим физическим и химическим свойствам от равновесных. [2]
В молекуле дезоксигемоглобина железо отстоит от плоскости порфиринового кольца примерно на 0,5-0,6 А° (есть небольшие отличия между a- и b-субъединицами). Из шести 3d электронов железа Fe(II) два электрона спарены на одной из низших d -орбиталей ( dxy , dyz , dxs ), а четыре электрона занимают оставшиеся d -орбитали, их спиновые моменты, согласно правилу Хунда, параллельны и суммарный спин S -2. Магнитный момент гема в этом состоянии равен ~ 5,5 боровского магнетона (БМ), а спектр поглощения в зеленой области имеет характерную полосу с lmax ~ 556 нм. Присоединение кислорода ведет к значительным изменениям. Атом железа в оксигемоглобине лежит практически в плоскости порфиринового кольца (расстояние до плоскости составляет 0,16 А° в α- и 0,00 А° в β- субъединицах). Все шесть d -электронов спарены на трех низших d -орбиталях, S= 0, оксигемоглобин диамагнитен. В зеленой области спектра имеются две характерные полосы поглощения: а (lmax 576 А°) и b (542 ??).
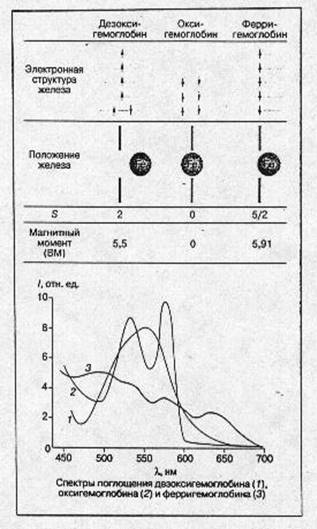
Рис.4. Основные характеристики молекулы гемоглобина в различных состояниях.
В ферригемоглобине (метгемоглобин) при нейтральных значениях рН место кислорода занимает молекула воды (при щелочных значениях рН-ОН), железо находится значительно ближе к плоскости гема, чем в дезоксигемоглобине, все пять d -электронов неспарены и занимают пять d- орбиталей. S = 5/2 и магнитный момент равен 5,91 БМ.
Структурные изменения в активном центре (вблизи гема) приводят и к значительным изменениям пространственной структуры всего белка. При оксигенации (переход от Т- к R-форме) смещение отдельных аминокислотных остатков достигает 7 А°. Как уже было сказано выше, четвертичная структура гемоглобина характеризуется наличием четырех полипептидных цепей, образующих две (a- и две b-субъединицы). Более детальные исследования показали, что субъединицы образуют ab-димеры. ТÞR – переход сопровождается поворотом одного димера относительно другого на 12-15° и в конечном счете приводит к увеличению карманов, в которых находятся гемы. Эти структурные изменения инициируются присоединением первой молекулы О2 к одному из свободных гемов и распространяются на всю глобулу. Именно поэтому в равновесной смеси всегда присутствуют только Т- и R-формы. Эти димеры в Т-форме стягиваются 14 дополнительными (по сравнению с R-формой) солевыми мостиками (водородные связи между ионными или нейтральными группами аминоксилот, ван-дер-ваальсовы контакты). Кроме того, между b-субъединицами в Т-форме присоединяется молекула дифосфоглицерата, что также приводит к сужению карманов. Эти изменения схематически представлены на рис.5.
Триггером для всех описанных выше структурных перестроек при переходах между Т- и R-формами и обратно служит присоединение или отщепление кислорода. После локального элементарного химического акта: присоединение или отщепление низкомолекулярного лиганда, окисление железа при образовании ферригемоглобина (иначе говоря, после появлении лишнего положительного заряда на железе) – возникает существенно неравновесное конформационное состояние – изменения вблизи активного центра уже произошли, а вся огромная молекула белка осталась в прежнем, еще не отрелаксировавшем состоянии. Последующая релаксация может занимать микросекунды ?