Реферат: Компьютерное моделирование в экологии
Как уже было сказано, скорость роста зависит от концентрации биогенных веществ. Для фитопланктона элементами, способными лимитировать рост, могут являться, например, азот, фосфор и углерод. Возможные способы отражения этого факта в работе С.Йоргенсена описаны следующим образом
![]() ,
,
где P, N, C – концентрация растворенного фосфора, азота и углерода; KP , KN , KС – соответствующие константы полунасыщения. Возможно также использование средней величины лимитирующих факторов: 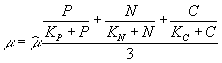 .
.
В работах Н.С.Абросова с коллегами была предложена следующая модель динамики экосистемы. Рассматривают организмы одного трофического уровня, которые существуют за счет совместного потребления ресурсов питания. Пусть ij (s1 ,...,sm ) – скорость потребления единицей биомассы (особью) вида i ресурса j ; i = i (s1 ,...,sm ) – удельная скорость прироста биомассы (плотности) i -го вида; s = (s1 ,...,sm ) – вектор концентрации ресурсов в среде; di – удельная скорость элиминации организмов i -го вида; Аj – скорость поступления j -го субстрата извне на единицу объема экосистемы; Di – удельная скорость удаления ресурса в открытой системе. Тогда динамика системы описывается уравнениями
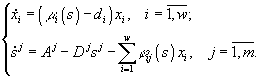 (1)
(1)
Модель, описывающая культивируемое в хемостате сообщество, является частным случаем модели (3.1) при условиях di = Dj = D , ![]() = Aj /D , где D – скорость разведения культуры,
= Aj /D , где D – скорость разведения культуры, ![]() – концентрация j -го субстрата в поступающей в реактор питательной среде, и задается системой уравнений
– концентрация j -го субстрата в поступающей в реактор питательной среде, и задается системой уравнений
![]()
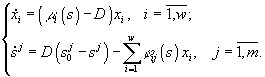
(Хемостат – непрерывный культиватор, в котором за счет саморегулирующх механизмов популяции поддерживается постоянная плотность популяции.)
В случае взаимозаменяемых ресурсов зависимость i (s ) описывается выражением
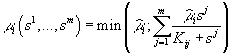
или его линейной аппроксимацией
![]() ,
,
где ij – элементы матрицы приспособленности: ![]() , здесь ij = cij Yij , где cij – удельная скорость потребления j -го субстрата i -м видом организма, а Yij – экономический коэффициент использования j -го вещества i -м видом.
, здесь ij = cij Yij , где cij – удельная скорость потребления j -го субстрата i -м видом организма, а Yij – экономический коэффициент использования j -го вещества i -м видом.
Если ресурсы взаимонезаменяемы, то
![]()
или
![]() .
.
В приведенных выше формулах символ ![]() обозначает максимально возможную удельную скорость роста организмов i -го вида; Kij – константа полунасыщения роста i -го вида на j -м субстрате.
обозначает максимально возможную удельную скорость роста организмов i -го вида; Kij – константа полунасыщения роста i -го вида на j -м субстрате.
В моделях, использующих дифференциальные уравнения, возможен учет лимитирования роста световой энергией. В этом случае предлагается использовать следующую зависимость:
![]()
здесь xi – концентрация клеток i -го вида; μi – удельная скорость роста i -го вида; ![]() – максимально возможная удельная скорость роста организмов i -го вида;
– максимально возможная удельная скорость роста организмов i -го вида; ![]() – процентное содержание хлорофилла в биомассе; li – коэффициент поглощения света i -м видом;
– процентное содержание хлорофилла в биомассе; li – коэффициент поглощения света i -м видом; ![]() – КПД фотосинтеза i -го вида клеток; Yi – величина, обратная калорийности биомассы i -го вида клеток; E – мощность ФАР (фотосинтетически активной радиации), падающей на поверхность культуры; d – толщина слоя культуры в кювете культиватора (приведено выражение для удельной скорости μi в случае лимитирования совместного роста двух видов только энергией света) (Абросов, Ковров, 1977).
– КПД фотосинтеза i -го вида клеток; Yi – величина, обратная калорийности биомассы i -го вида клеток; E – мощность ФАР (фотосинтетически активной радиации), падающей на поверхность культуры; d – толщина слоя культуры в кювете культиватора (приведено выражение для удельной скорости μi в случае лимитирования совместного роста двух видов только энергией света) (Абросов, Ковров, 1977).
А.Я.Болсуновским (1999) была предложена следующая математическая модель динамики компонентов сообщества из двух видов (спирулина и хлорелла), рост которых лимитируется интенсивностью света всей области ФАР

где xi – концентрация клеточного хлорофилла; μi – удельные скорости роста; ai , bi – константы уравнения Михаэлиса-Ментен; ![]() – средняя освещенность для всего сообщества; E0 – поверхностная освещенность; D – скорость протока.
– средняя освещенность для всего сообщества; E0 – поверхностная освещенность; D – скорость протока.
Кроме того, в моделях возможно отразить эффекты метаболического воздействия, как, например, это было сделано Ю.А.Домбровским с соавторами (1990). Скорость роста фитопланктона i -го вида описывалась выражением
![]() ,
,
где x1 , x2 , s – концентрации двух видов фитопланктона и минерального вещества, выраженные в единицах лимитирующего биогенного вещества; ![]() – коэффициенты метаболизма; Mi – максимальная скорость фотосинтеза; Hi – параметр насыщения;
– коэффициенты метаболизма; Mi – максимальная скорость фотосинтеза; Hi – параметр насыщения; ![]() – эмпирический коэффициент ингибирования i -го вида j -м.
– эмпирический коэффициент ингибирования i -го вида j -м.
Как уже указывалось выше, при моделировании динамики биомассы фитопланктона, чаще всего используют уравнение Моно, описывающее зависимость удельной скорости роста популяции μ от концентрации s лимитирующего рост субстрата
![]() , (2)
, (2)