Реферат: Координація періодичної діяльності гастродуоденальної зони при порушенні її функціонально-морфологічної цілісності
Методи дослідження: використані електрофізіологічні методи дослідження міоелектричної активності шлунка та дванадцятипалої кишки, методи збору та біохімічного аналізу шлункового соку, гістохімічні методи оцінки стану слизової оболонки шлунка та дванадцятипалої кишки, варіаційно-статистичні методи обробки експериментальних даних: розрахунок середньої та її похибки.
Мета дослідження: встановлення взаємозв’язків періодичної моторної активності шлунка, дванадцятипалої кишки, діяльності секреторних залоз шлунка в умовах порушення морфофункціональної цілісності гастродуоденальної зони.
Завдання наукового дослідження:
1) дослідити стан моторної активності шлунка при моделюванні вегетативного дисбалансу;
2) дослідити стан секреторного апарату шлунка при моделюванні вегетативного дисбалансу;
3) вивчити міоелектричну активність шлунка та дванадцятипалої кишки при моделюванні дуоденогастрального рефлюксу;
4) з’ясувати стан секреторної функції шлунка при моделюванні дуоденогастрального рефлюксу;
5) зіставити особливості моторної та секреторної функцій верхніх відділів шлунково-кишкового тракту при моделюванні дуоденогастрального рефлюксу;
6) дослідити стан моторної діяльності шлунка та дванадцятипалої кишки, а також секреторної активності шлунка при блокаді місцевих нітрергічних механізмів в інтактних тварин та при моделюванні дуоденогастрального рефлюксу;
7) з’ясувати участь нітрергічних механізмів у регуляції періодичної моторно-секреторної діяльності верхніх відділів шлунково-кишкового тракту в інтактному стані та при моделюванні дуоденогастрального рефлюксу.
Наукова новизна одержаних результатів. У роботі вперше проведено комплексне експериментальне дослідження моторно-секреторної діяльності гастродуоденальної зони на первинних етапах розвитку вегетативного дисбалансу та дуоденогастрального рефлюксу. Уперше встановлено, що при розвитку вегетативного дисбалансу зміни моторно-секреторних ритмів шлунка протікали в три стадії, що супроводжувалось зниженням функціональної здатності секреторних залоз шлунка, явищем «відмови в регуляції», якому передувала поява кислої шлункової секреції натще, а також розвитком дуоденогастрального рефлюксу.
Уперше показано, що на первинних етапах розвитку дуоденогастрального рефлюксу результатом адаптивно-компенсаторних перебудов періодичної діяльності гастродуоденальної зони було підвищення активності парієтальних та поверхнево-епітеліальних клітин слизової оболонки шлунка із супутнім пригніченням діяльності її головних клітин. Уперше виявлено, що при розвитку дуоденогастрального рефлюксу нітрергічна регуляція зберігає свій гальмівний вплив у відношенні скоротливої активності гастродуоденальної зони шляхом пригнічення міогенної пікової активності антрального відділу шлунка та цибулини дванадцятипалої кишки. При цьому відбувається перебудова місцевої нейроендокринної регуляції гастродуоденальної зони, однією з механізмів якої є протилежна зміна впливу NO-ергічної ланки у напряму діяльності парієтальних клітин шлунка. Новими є дані, які свідчать, що тривале введення блокаторів NO-синтази може призводити до порушень цілісності слизової оболонки шлунка та виникнення виразкових ушкоджень останньої.
Практичне значення одержаних результатів. Отримані в роботі дані можуть бути використані в науково-дослідній роботі при вивченні клініко-фізіологічних аспектів загального адаптаційного синдрому за умов розвитку стрес-реакції. Уперше запропоновано оригінальний нетравматичний спосіб збору шлункового соку у лабораторних щурів. Результати даної роботи можна враховувати в клінічній гастроентерології при вивченні змін періодичної моторно-секреторної діяльності верхніх відділів шлунково-кишкового тракту при розвиткові дуоденогастроезофагеальної рефлюксної хвороби. Також основні положення роботи можуть бути врахованими при розробці нових функціональних проб та діагностичних критеріїв розвитку функціонально-морфологічних зсувів гастродуоденальної зони на перших етапах передонкологічних процесів у шлунці.
Практико-теоретична цінність дисертаційного дослідження підтверджується впровадженням результатів та пропозицій, які витікають з роботи, в навчально-практичний процес кафедри фізіології людини і тварин Дніпропетровського національного університету та в науково-дослідницьку діяльність ДУ “Інституту гастроентерології АМН України”.
Особистий внесок здобувача. Автором самостійно: проведено аналіз літературних джерел, виконані експерименти з вивчення міоелектричної і секреторної активності шлунка та дванадцятипалої кишки у щурів; запропоновано та обґрунтовано оригінальний метод збору шлункового соку у щурів; проведено систематизацію та статистичний аналіз показників моторики та секреції гастродуоденальної зони завдяки відомим комп’ютерним програмам. У дисертації не використовувалися практичні розробки співавторів опублікованих робіт. У працях, написаних у співавторстві, здобувачеві належить: проведення (або участь у проведенні) експериментів, аналіз, систематизація та узагальнення результатів дослідження, та оформлення їх у вигляді статей.
Апробація результатів. Матеріали дисертації представлені у доповідях на конференціях ДНУ за підсумками науково-дослідних робіт у 2005–2007 рр.; на IV Міжнародній науково-практичній конференції “Динаміка наукових досліджень – 2005” (Дніпропетровськ, 2005 р.); на Міжнародній науковій конференції, приуроченій до 60-ліття новоствореної кафедри фізіології людини і тварин Львівського університету ім. І. Франка (Львів, 2006 р.); на Всеукраїнській науково-практичній конференції «Вклад молодих вчених в розвиток медичної науки і практики» (Харків, 2006 р.); на VIII Міжнародній науково-практичній конференції «Людина і космос. Ракетно-космічна техніка» (Дніпропетровськ, 2006 р.); на ІІ Міжнародній конференції “Neural-humoral and cellular regulatory mechanisms of digestion processes” (Київ, 2005 р.); на XVII З’їзді українського фізіологічного товариства з міжнародною участю (Чернівці, 2006 р.); на IХ Міжнародній науково-практичній конференції «Людина і космос. 50-ліття космічної ери» (Дніпропетровськ, 2007 р.); на Всеукраїнській науковій конференції «Сучасні питання фізіології та медицини», присвяченій 85-річчю заснування кафедри фізіології людини і тварин та 100-річчю з дня народження П. Є. Моцного (Дніпропетровськ, 2007 р.).
Публікації. За темою дисертації опубліковано 10 робіт: 4 статті – у фахових журналах ВАК, 6 є тезами доповідей з’їздів і конференцій.
Структура та обсяг дисертації. Дисертація складається зі вступу, огляду літератури, опису методів досліджень, двох розділів результатів досліджень, узагальнення результатів досліджень, висновків і списку використаних джерел. Дисертація викладена на 157 сторінках, містить 55 рисунків, 2 таблиці, список використаних джерел із 280 найменувань.
ОСНОВНИЙ ЗМІСТ
Методи дослідження. Всі експерименти були проведені відповідно до існуючих міжнародних правил використання лабораторних тварин у медико-біологічних дослідженнях. Для вирішення встановлених задач було проведено 6 серій експериментів (гострих, напівгострих та хронічних). У першій серії експериментів (N=15) вивчали міоелектричну активність шлунка собак у вихідному стані та в динаміці моделювання вегетативного дисбалансу. У другій серії (N=5) досліджували секреторну активність шлунка собак у вихідному стані та в динаміці моделювання вегетативного дисбалансу. У третій серії (N=22) досліджень вивчали міоелектричну активність шлунка та дванадцятипалої кишки інтактних щурів у вихідному стані та при блокуванні NO-синтази. У четвертій серії (N=24) досліджень вивчали секреторну функцію шлунка інтактних щурів у вихідному стані та при блокуванні NO-синтази. У п’ятій серії (N=27) вивчали міоелектричну активність шлунка та дванадцятипалої кишки при моделюванні дуоденогастрального рефлюксу відповідно на 3-й, 6-й та 12-й день у вихідному стані та при блокуванні NO-синтази. У шостій серії (N=36) досліджували шлункову секрецію у щурів при моделюванні дуоденогастрального рефлюксу відповідно на 3-й, 6-й та 12-й день у вихідному стані та за умов неселективного блокування NO-синтази. Моделювання вегетативного дисбалансу у собак проводили шляхом щоденних внутрішньом’язових ін’єкцій карбохоліну (0,01 мг/кг) за 15 хвилин до введення цинхофену (0,05 г/кг) натще per os протягом 30 діб. Для відтворення дуоденогастрального рефлюксу здійснювали щоденне введення жовчі per os натще із наступним впровадженням імобілізаційно-холодового стресу протягом однієї години (+4˚С) [Тарасенко и др., 2001, з модифік.]. Міоелектричну активність шлунка та дванадцятипалої кишки вивчали за допомогою платинових (у собак) та срібних (у щурів) біполярних електродів з використанням самописця WI-387G та системи «поліграф RM-86 Nihon Kohden–комп’ютер». Міоелектричну активність оцінювали за характером змін її складових (частоти та амплітуди повільних потенціалів, періоду основного (базального) електричного ритму, а також тривалості фаз мігруючого моторного комплексу). Неселективне блокування NO-синтази здійснювали за допомогою внутрішньочеревної ін’єкції NG-нітро-L-аргініну (L-NNA). Для вивчення секреторної функції шлунка собакам вживляли фістули за В. А. Басовим. Шлунковий сік у щурів збирали зондовим методом. Для з’ясування стану слизової оболонки гастродуоденальної зони здійснювали макроскопічні та гістохімічні дослідження відповідних ділянок слизової оболонки шлунка та дванадцятипалої кишки із забарвленням гематоксилін-еозином, альциановим синім, та ШИК-реакцію за загальноприйнятими методиками [Крышень и др., 1978]. Активність NO-синтаз в секреторних залозах шлунка визначали за допомогою NADPH-діафоразної реакції [Shyamal et al., 2001]. Статистичну обробку отриманих результатів проводили за допомогою пакету програм Microsoft Excel та Origin 6.1 Pro з використанням t-критерію Стьюдента, а також непараметричних критеріїв Вілкоксона-Манна-Уітні для порівняння рядів даних з малою кількістю значень (N) [Гланц, 1999]. Досліджувані співвідношення вважали достовірними при Р<0,05.
Результати дослідження. Міоелектрична активність шлунка при моделюванні вегетативного дисбалансу. Моделювання вегетативного дисбалансу супроводжувалося розвитком виразкових ушкоджень слизової оболонки шлунка, які розташовувалися в місці переходу інтрамедіальної зони шлунка в препілоричну, або ж на ділянці воротаря на малій кривині шлунка. Якщо в інтактному стані міоелектрична активність шлунка складалася з 3-х фаз: фази I (60-90 хв), фази II (20–30 хв) і фази III (5-7 хв), то на 3–5-й день експерименту спостерігалося збільшення тривалості II-III фаз на 150,0–200,0% (до 40 хв), а тривалість I фази зменшилася на 30,0–40,0% (до 40–55 хв, P<0,01). Треба зазначити, що на 7–10-й день дослідження можна було спостерігати розповсюдження повільних хвиль міоелектричної активності в ретроградному напрямку, що передувало закиданню дуоденального вмісту в шлунок. У подальшому – на 15–35-й день – такий стан моторної активності супроводжувався антиперистальтичними хвилями, які часто закінчувалися блювотою. На ранніх етапах моделювання вегетативного дисбалансу відзначалася тенденція до зростання частоти повільних хвиль основного електричного ритму шлунка – до 10-го дня (рис. 1, А), при цьомуперіод основного електричного ритму зменшувався (P<0,05).
А 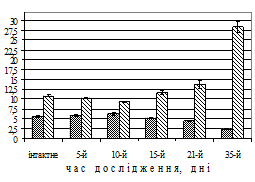 | Б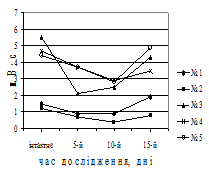 |
Рис. 1. Динаміка частоти (темні стовпчики) повільних потенціалів, періоду основного електричного ритму (світлі стовпчики) антрального відділу шлунка (А) та моторного індексу в ряді відведень шлунка собак (Б) при моделюванні вегетативного дисбалансу (зліва на рис. Б – схема розташування та номери відведень). По вісі абсцис – час від початку досліду, день; по вісі ординат – частота повільних потенціалів, за хв, період основного електричного ритму, с, значення моторного індексу, мВ∙с (M±m (А), M (Б), N=15 (інтактне), N=5 (дослід)).
У подальшому – на 15-й день – навпаки, зменшилася частота повільних потенціалів та зріс період основного електричного ритму майже в 3 рази (P<0,01). Треба зазначити, що вже з 3–5-го дня відзначалося зниження моторного індексу, особливо в перехідній зоні між антральним та фундальним відділами шлунка (рис. 1, Б). Неузгодженість у діяльності між фундальним та антральним відділами шлунка за даними моторного індексу можна було виявити саме до 15–20-го дня дослідження.
Секреторна діяльність шлунка при моделюванні вегетативного дисбалансу. Дослідження другої серії виявили, що вже на 10-й день у більшості тварин поряд із підвищенням слизевиділення натще спостерігалося виділення кислого соку (рис. 2, А). При цьому тривалість та інтенсивність останнього мали індивідуальні особливості у кожної тварини. У шлунковому вмісті визначався пепсин, максимальна кількість якого припала на 21-й день. Починаючи з 10-го дня можна
А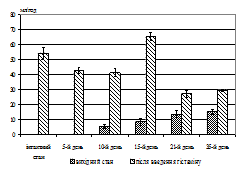 | Б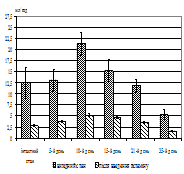 |
Рис. 2. Зміни дебіту базальної кислої шлункової секреції, об’єму кислої шлункової секреції (А) та дебіту слизу (Б) у собак після введення гістаміну в інтактному стані та в динаміці вегетативного дисбалансу. По вісі абсцис – час від початку досліду, дні; по вісі ординат – дебіт об’єму, мл/год (M±m, N=15 (інтактний стан), N=5 (дослід)).
було спостерігати закидання дуоденального вмісту в шлунок, про що свідчила наявність жовчних кислот, концентрація яких досягала максимальних величин наприкінці експерименту. При цьому рН шлункового соку змінювався відповідно та на 15-й день дослідження досяг мінімальних значень, що вже не змінювалися до кінця експерименту. Загалом на 7–10-й день у міжтравний період з’явилася кисла шлункова секреція натще, збільшилося виділення слизу й у шлунковому вмісті виявлялися жовчні кислоти. Для перевірки причин появи кислої секреції застосовували атропіновий тест, який був позитивним. У той же час, поява жовчних кислот у шлунковому вмісті обумовлена закиданням дуоденального вмісту в шлунок. Причиною цього дуоденогастрального рефлюксу, у свою чергу, є антиперистальтичні скорочення гладеньком’язової мускулатури дванадцятипалої кишки, що пов’язане із включенням компенсаторних механізмів, спрямованих на ліквідацію закислення в гастродуоденальній зоні. Перевірка функціональної здатності секреторних залоз шлунка за допомогою гістамінового тесту виявила також зміни резервних можливостей секреторного апарату шлунка (рис. 2, А, Б).
Міоелектрична активність шлунка та дванадцятипалої кишки щурів в інтактному стані. В інтактному стані міоелектрична активність гастродуоденальної зони мала фазний характер, при цьому фаза спокою становила 40-50 хв, фаза нерегулярної пікової активності – 50-60 хв, фаза регулярної пікової активності – 5-9 хв. В інтактному стані період основного електричного ритму шлунка склав 11,88±0,20 с (P<0,05), а дванадцятипалої кишки – відповідно 1,61±0,02 с (P<0,05). Однак у ряді експериментів період основного електричного ритму шлунка не перевищував 8,30±0,34 с (P>0,05), а в інших тварин в окремих експериментах цей показник дванадцятипалої кишки досяг 2,10±0,15 с (P<0,05), що вказує на індивідуальні особливості моторики верхніх відділів шлунково-кишкового тракту.
Міоелектрична активність шлунка та дванадцятипалої кишки, та секреторна діяльність шлунка в інтактних тварин за умов блокування NO-синтази. В інтактних тварин уже через 5–7 хв після введення блокатору NO-синтази в складі міоелектричної активності спостерігалося збільшення амплітуди маскуючих коливань, на тлі яких почали проявлятись поодинокі пікові потенціали, а через 35–45 хв спостерігали електричні коливання стінки шлунка, які були схожі за складом із ІІ-ІІІ фазою міоелектричної активності; така картина зберігалася протягом наступних 75–85 хв. При цьому електричні коливання, які були найбільш подібні до ІІІ фази, спостерігалися в шлунці через 80–100 хв та тривали не більше 10–15 хв. При аналізі міоелектричної активності дванадцятипалої кишки при блокаді нітрергічних механізмів відзначено збільшення амплітуди основного електричного ритму за рахунок підвищення амплітуди першого висхідного коліна повільної хвилі вже через 12–16 хв після введення L-NNA, а через 20 хв спостерігали появу ІІІ фази міоелектричної активності. Треба зазначити, що як у вихідному стані, так і після введення L-NNA, міоелектрична активність дванадцятипалої кишки містила нестійкі за появою повільнохвильові коливання, період яких, як правило, наближався за значенням до періоду основного електричного ритму шлунка та становив 6,80±0,15с. Значення основних показників міоелектричної активності шлунка та дванадцятипалої кишки після введення L-NNA на різних етапах розвитку дуоденогастрального рефлюксу наведені в таблиці 1.
Таблиця 1.
Основні показники міоелектричної активності шлунка та дванадцятипалої кишки після введення L-NNA після 3-х, 6-ти, 12-ти днів моделювання дуоденогастрального рефлюксу (M±m)
| Показник | Інтактне (N=12) | Після 3-х днів ДГР (N=9) | Після 6-ти днів ДГР (N=9) | Після 12-ти днів ДГР (N=9) |
| Період ОЕР шлунка | 10,85±0,26* | 11,16±0,39* | 14,66±0,57 | 15,79±0,68 |
| Період ОЕР ДПК | 1,60±0,02 | 1,73±0,03 | 1,96±0,04 | 1,93±0,06** |
| Амплітуда ОЕР шлунка | 40,80±4,08 | 15,12±2,20 | 25,88±3,05 | 13,96±2,52** |
| Амплітуда ОЕР ДПК | 24,12±1,64* | 26,10±1,37 | 54,15±2,97* | 31,28±1,39 |
| Моторний індекс шлунка | 8,25±1,06 | 4,49±1,35 | 7,59±1,03 | 5,99±1,19** |
| Моторний індекс ДПК | 6,68±0,98 | 7,76±1,74 | 13,50±0,67 | 8,88±0,64 |