Реферат: Механізм дії високого тиску і температури на деякі мікроорганізми та вітаміни
Кінетику інактивації мікроорганізмів і денатурації білків під дією сталих підвищеної температури й тиску можна розглядати як незворотну хімічну реакцію першого порядку. Зміна концентрації мікроорганізмів, що вижили, і не денатурованих білків в залежності від часу C(t) описується рівнянням розпаду:
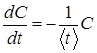 , (1)
, (1)
рішенням якого є: 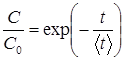 (2)
(2)
де ![]() – імовірність переходу біомолекули або мікроорганізму в пошкоджену форму в одиницю часу. У відповідності до теорії активованого комплексу, розглядаючи вказаний перехід як одновимірний процес активаційного типу, маємо:
– імовірність переходу біомолекули або мікроорганізму в пошкоджену форму в одиницю часу. У відповідності до теорії активованого комплексу, розглядаючи вказаний перехід як одновимірний процес активаційного типу, маємо:
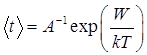 , (3)
, (3)
де W– висота енергетичного бар’єру, що відокремлює нативний та денатурований стани. Висота вказаного енергетичного бар’єру дорівнює мінімальній роботі, яка затрачується при переході з нативного стану на вершину енергетичного бар’єру, тобто:
![]() , (4)
, (4)
де Р0 і Т0 – температура і тиск у барокамері, ΔE, ΔS, ΔV– зміна енергії, ентропії та об’єму зразка.
При ΔV<0 вільна енергія реакції зменшується з ростом тиску, що відповідає зрушенню хімічної рівноваги убік збільшення кінцевого продукту у повній відповідності із принципом Ле-Шательє. Таким чином, зовнішній тиск P впливає на рівноважне співвідношення концентрацій у нативному N і денатурованому D станах через доданок Р0 ΔV у термодинамічному потенціалі, а також впливає на швидкість денатурації через зміну активаційного бар'єра (U + Р0 ΔV, де U= ΔE– Т0 ΔS).
Рішенням системи рівнянь (2)-(4) методом найменших квадратів винайдено набір {ln(A), U й ΔV}, при якому теоретичні криві щонайкраще проходять через всі експериментальні крапки. Цінність цього рішення полягає в тому, що за допомогою всього лише трьох параметрів описана реакція системи на будь-яке сполучення зовнішніх впливів тиску P, температури T й тривалості їхньої дії t. Непрямим підтвердженням правильності отриманих параметрів є близькість за порядком величини U та ΔV МАФАМ і цвілі до відомих значень для різних білків. Отримана зміна молярного об'єму ΔV для невеликої молекули вітаміну С по модулю на порядок менша, ніж характерні значення цього параметру для мікроорганізмів або для білків.
На Рис. 1 представлено криві, що розраховані з використанням винайдених параметрів, в порівнянні з експериментальними даними. Теорія цілком задовільно описує експеримент, зокрема, криві передають правильний хід концентрації залежно від тиску як для вітаміну С, так і для МАФАМ і цвілі, а також правильно відображають тенденції зміни цих залежностей при зростанні температури й часу обробки. У рамках однієї й тієї ж формули утворюються як плавно спадаючі майже лінійні залежності для вітаміну С, так і східчасті криві для МАФАМ і цвілі.
Тим самим проведені розрахунки, з одного боку, підтверджують правильність обраної моделі, а з іншого – дають можливість планувати експеримент і вибирати оптимальні значення параметрів обробки для максимальної стерилізації продуктів при найкращій збереженості вітаміну С.
Досліджено вплив високого тиску на кількість мікроорганізмів групи МАФАМ у яблучному пюре, тобто знайдені значення параметрів ln(A·1сек)=13.2, U=13.4ккал/моль та ΔV=-25.1мл/моль, які є близькими до значень відповідних величин, що отримані для вишневого соку. На Рис. 3 представлені експериментальні й теоретичні значення відносних чисел КУО МАФАМ (з двох серій дослідів T=30°С, t=20хв і T=25°С, t=10хв), отриманих для яблучного пюре, а також криві, що розраховані для вишневого соку у тих же умовах. Має місце цілком задовільна якісна згода теорії з експериментом. Крім того, в обох випадках наявне помітне зрушення убік менших тисків (приблизно на 150МПа) кривих для яблучного пюре порівняно з кривими для вишневого соку. Таке зрушення кривих убік менших тисків обумовлюється різними значеннями кислотності в яблучному пюре (pН=5.3) і у вишневому соку (pН=6.1). Як відомо, процеси денатурації білків й виживаність мікроорганізмів визначаються, насамперед, кислотністю середовища, швидкість інактивації мікроорганізмів тиском росте зі зниженням рН.
Для з’ясування, наскільки широким є клас об'єктів, для яких справедлива використана модель, та чи буде вона описувати відомі дані з інактивації мікроорганізмів, що отримані в роботах інших авторів, запропонований підхід було застосовано щодо даних з інактивації спор Bacillus subtilis і Bacillus stearothermophilus з робіт Urzica A.C., 2004 р. та Ardia A., 2004 р. Отримані таким чином значення термодинамічних параметрів наведені в Табл. 1. Для порівняння там же наведені знайдені нами параметри інактивації вегетативних форм мікроорганізмів.
Як видно, значення активаційного бар'єру U (Табл. 1) є близькими для всіх об'єктів. Крім того, ці значення потрапляють в інтервал від 10 до 20ккал/моль, характерний для активаційного бар'єру процесу денатурації різних білків. Отже, можна припустити що це є одна і та ж величина і що бар'єр інактивації U різних мікроорганізмів визначається процесом, що не пов'язаний з їхнім розміром, масою або формою.
Значення зміни питомого об'єму ΔV для всіх об'єктів (Табл. 1) за порядком величини збігаються з характерними значеннями ΔV для білків. Таким чином підтверджується припущення, що інактивація мікроорганізмів обумовлена деградацією білків. Величини ΔV для різних вегетативних форм приблизно однакові. Теж саме маємо і для спор, при цьому ΔV для спор помітно менше, ніж для вегетативних форм. Цей факт також підтверджує зв'язок інактивації мікроорганізмів з деградацією білків, оскільки в спорах упаковка білків є більш щільною і в них відсутні або є в мінімальній кількості порожнини, які заповнюються водою при розгортанні білка і дають вагомий внесок у зміну об’єму зразка.
Отримані в роботі експериментальні дані досить добре описуються в рамках кінетичної моделі першого порядку, одначе, в літературі часто зустрічаються дані зі значними відхиленнями від лінійності, які зазвичай описуються комбінацією двох реакцій першого порядку як двофазна кінетика з різними швидкостями інактивації. Зміна порядку реакції іноді спостерігається у межах одного виду бактерій і навіть у межах однієї серії дослідів, що зазвичай пояснюється існуванням невеликої частини популяції з підвищеним опором до впливу високого тиску, «багатофазністю» процесу інактивації або зміною порядку реакції. В дисертації показано, що цей ефект є пов'язаним з нерівномірним остиганням зразку, який нагрівається внаслідок стискання. При адіабатичному стисканні під тиском до 600МПа константа швидкості інактивації ![]() може збільшуватися для різних об’єктів від 2 до 5 разів у залежності від співвідношення W/kT.
може збільшуватися для різних об’єктів від 2 до 5 разів у залежності від співвідношення W/kT.
Вирівнювання температури у зразку, що стискається, описується нестаціонарним рівнянням теплопровідності. Аналіз показує, що характерний час експоненціального остигання τ залежить від конструкції барокамери та не залежить від ступеню нагріву. Оцінка часу остигання дає τ ~10хв, що влучає в область звичайних експозицій (від 5 до 30хв). Таким чином, при знятті кінетичної кривої дослідники неминуче охоплюють як область t < τ, так й t > τ. При t < τ температура біля середини зразка вища, ніж на периферії. При t > τ температура вирівнюється, середня по об'єму швидкість реакції падає, що й спостерігається як нова «фаза з підвищеною опірністю тиску». Можливість фіксації цієї фази у досліді залежить, насамперед, від величини зміни температури, яка визначається як величиною самого тиску, так і часом його зростання порівняно з τ.
Урахування нерівномірного остигання стиснутого зразка здійснювалось через усереднення по об'єму рішення лінійного кінетичного рівняння, в якому константа швидкості реакції ![]() визначається за формулою Ареніуса (3) з неоднорідним розподілом температури
визначається за формулою Ареніуса (3) з неоднорідним розподілом температури
![]() ,(5)
,(5)
де ![]() – рішення нестаціонарного рівняння теплопровідності, безрозмірний розподіл температури в одиницях її максимального перевищення над початковою, що задається величиною адіабатичного нагрівання при стисненні βP (для води β=3ºC/100МПа). Початок відліку часу t=0 відповідає закінченню підвищення тиску.
– рішення нестаціонарного рівняння теплопровідності, безрозмірний розподіл температури в одиницях її максимального перевищення над початковою, що задається величиною адіабатичного нагрівання при стисненні βP (для води β=3ºC/100МПа). Початок відліку часу t=0 відповідає закінченню підвищення тиску.
При урахуванні нерівномірного остигання криві залежності ступеню деградації біооб’єктів від тиску (Рис. 1) змінюються незначно. Граничне значення тиску, при якому спостерігається різкий спад концентрації, зрушується в сторону менших тисків на 20 і 10МПа для t=10 й 20хв відповідно. З ростом часу обробки виправлення на нагрівання зменшується за рахунок зменшення середньої температури, при якій відбувається інактивація. Ґрунтуючись на отриманих результатах у роботі відкинуто сумнівні припущення, які робляться для пояснення особливостей кінетики цих процесів. З іншого боку, той факт, що лінійна кінетична модель здатна описати спостережувані на експерименті відхилення від лінійності є підтвердженням адекватності цієї моделі.
Розглянуто зростання кислотності під тиском як фактор інактивації мікроорганізмів. Аналіз літератури показує, що концентрація іонів водню визначає межі існування живої матерії. Багато експериментальних робіт доводять, що кислотність харчового середовища істотно впливає на ступінь інактивації мікроорганізмів високим тиском. Отримані і співставленні у даній роботі експериментальні і розрахункові криві для яблучного пюре і вишневого соку (Рис. 3) підтверджують факт росту швидкості інактивації мікроорганізмів під тиском за знижених рН. Величина зрушення збігається з літературними даними, згідно з якими кислотність харчових продуктів збільшується під дією тиску від 0.2 до 0.5 одиниць рН на кожні 100МПа. На відміну від мікроорганізмів вітамін С стабілізується при підвищенні кислотності середовища. Результати вимірів впливу тиску на зміст вітаміну С у середовищах різної кислотності (Рис. 2) показують, що більша деградація вітаміну С спостерігається у розчинах з більшим рН. Той же ефект дає збільшення часу обробки. Зростання кислотності харчових продуктів з підвищенням тиску пояснюється добре вивченим ростом дисоціації молекул води при підвищенні тиску й температури. В роботі показано, що збільшення тиску на 100МПа зменшує pH приблизно також, як збільшення температури на 10ºC. Таким чином збіг змін показника pH в інтервалах температури 60-100ºC та тиску 500-1000МПа, які стерилізують продукти харчування, не є випадковим і саме зміна pH з ростом як температури, так і тиску приводить до денатурації білків і, відповідно, до інактивації мікроорганізмів. Інактивуюча дія високого тиску зумовлена, в основному, зміною рН середовища, а інактивація клітини є вторинним ефектом.
Спираючись на отримані результати доведено, що механізмом впливу тиску на мікроорганізми є денатурація білків. Денатурація білків при нагріванні й втрата ними біологічної активності пов'язані з розривом сітки з молекул води, яка обмежує або стримує вібраційну динаміку білка. Необхідна для розриву енергія визначає розмір бар'єру U для переходу з нативного в активований стан N → A# . Показником міцності сітки водневих зв'язків може служити кислотність середовища, оскільки, як доведено у роботі, вона впливає на швидкість інактивації. Величина рН знижується з ростом температури й тиску.
Вільна енергія реакції денатурації білка ΔGtotal формується (див. Рис. 5(а)) спільними ефектами згортання/розгортання та зміни ступеня гідратації неполярних і полярних груп білка. Ці внески значною мірою компенсують один одного, так що вільна енергія стабілізації типового білка становить усього 10-20ккал/моль, тоді як зміни ентальпії ΔHnonpolar й ΔHpolar (і помножені на температуру зміни ентропії TΔSnonpolar й TΔSpolar ) можуть розрізнятися більш, ніж на ±120ккал/моль. Там, де ΔGtotal <0, спостерігається теплова (при T>T2 ) і холодова (при T<T1 ) денатурація, що у залежності від умов експерименту може протікати швидко або дуже повільно. Область існування нативного стану T1 <T<T2 визначає температурний оптимум щодо активності білка.
На Рис. 5(б) проілюстровано залежність стабільності білка від тиску. Область тисків вище деякого критичного значення P>Pc відповідає денатурованому стану білка. Швидкість денатурації в цій області визначається величиною активаційного бар'єру, доданок U у якому пов’язаний з розривом сітки водневих зв'язків і є однаковим для термо- і бароденатурації. Вплив тиску на швидкість денатурації визначається сумарною зміною об'єму зразка. Область існування нативного стану P<Pc відповідає оптимуму активності білка під тиском, Pc є істотно меншим, ніж граничні значення інактивації (~500МПа для більшості мікроорганізмів).
Практичними наслідками дисертаційної роботи є можливості оптимізації параметрів стерилізації біооб’єктів та стандартизації обробки високим тиском. Запропонована модель дозволяє передбачити поводження досліджених об'єктів при різних сполученнях параметрів обробки: тиску, температури і тривалості їхньої дії. Як приклад у роботі наведено ряд номограм, розрахованих для вибору параметрів впливу на вишневий сік, що зменшує в 106 разів концентрацію МАФАМ і цвілі. Показано, що для порівнянного зменшення концентрації вітаміну С величина тиску повинна бути на порядок вище. Номограми призначені для вибору тиску P і температури T при заданих термінах обробки t або для вибору тиску P і терміну обробки t при заданих температурах T.
Наразі відсутні загальноприйняті критерії, що визначають якість обробки тиском харчових продуктів. Ґрунтуючись на отриманих результатах, в дисертації запропоновано використовувати усереднені термодинамічні характеристики мікрофлори ln(A), U й ΔV як контрольні параметри при розробці процедури стандартизації обробки продуктів високим тиском.
ВИСНОВКИ
У дисертаційній роботі експериментально досліджено закономірності та розроблена фізико-математична модель спільної дії високого тиску і температури на мікроорганізми і вітамін С у різних середовищах, вивчено мікроскопічний механізм впливу тиску на біосистеми. Отримані результати можуть бути використані для подальшого розвитку баробіології та нової технології стерилізації харчових продуктів високим тиском.