Реферат: Мембранные белки
Основная роль липидов в составе мембран заключается в стабилизации бислойной структуры, а белки являются активными компонентами биомембран. Мы обсудим некоторые принципы, оказавшиеся полезными для выяснения структурных особенностей мембранных белков. Мы приведем примеры, иллюстрирующие эти принципы.
На заре развития мембранологии полагали, что мембранные белки по своей структуре довольно гомогенны и уложены в виде 3-слоев по поверхности бислоя. Сейчас скорее склонны считать, что по крайней мере у трансмембранных белков те их участки, которые погружены в мембрану, содержат а-спирали. Конечно, очень хотелось бы сделать какие-то однозначные выводы по этому поводу, но они должны основываться на фактических данных. Перед лицом огромного структурного разнообразия растворимых белков приходишь к заключению, что интегральные мембранные белки могут оказаться гораздо сложнее, чем мы сейчас представляем. Классификация растворимых белков по типам структур была проведена только после того, как установили с высоким разрешением структуру более 100 различных белков. Что касается трансмембранных белков, то это удалось сделать только в одном случае — для белка фотосинтетического реакционного центра бактерий. Вместе с электронно-микроскопическими данными низкого разрешения о структуре бактериородопсина это единственный источник, на котором может основываться построение моделей для большинства других трансмембранных белков.
Еще один важный момент — способы прикрепления белков к мембране. Они схематически представлены на рис. 3.1.
1. Связывание с белками, погруженными в бислой. В качестве примеров можно привести Fi-часть Н + -АТРазы, которая связывает ся с Fo-частью, погруженной в мембрану; можно упомянуть также некоторые белки цитоскелета.
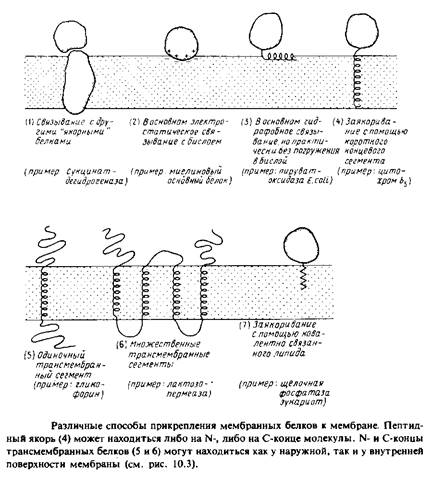
2. Связывание с поверхностью бислоя. Это взаимодействие имеет в первую очередь электростатическую природу или гидрофобную. На поверхности некоторых мембранных белков имеются гидрофобные домены, образующиеся благодаря особеностям вторичной или третичной структуры. Указанные поверхностные взаимодействия могут использоваться как дополнение к другим взаимодействиям, например к трансмембранному заякориванию.
3.Связывание с помощью гидрофобного «якоря»; эта структура обычно выявляется как последовательность неполярных аминокислотных остатков. Некоторые мембранные белки используют в качестве якоря кова-лентно связанные с ними жирные кислоты или фосфолипиды.
4.Трансмембранные белки. Одни из них пересекают мембрану только один раз, другие — несколько раз.
Различиями между наружными и внутренними мембранными белками не задается однозначно способ их прикрепления к бисЛою; эти различия определяют лишь относительную силу их связывания.
2. Очистка мембранных белков
Для очистки интегральных мембранных белков и получения их в биохимически активной форме необходимы детергенты, позволяющие солюбилизировать белки и сохранить их в растворе. Соответствующие требования к детергентам и правилам обращения с ними создают дополнительные проблемы помимо тех, с которыми обычно сталкиваются при очистке белков. Для выделения интегральных мембранных белков разработано много специальных методов, однако большинство схем очистки основано на тех же хроматографических и гидродинамических методиках, которые используются для растворимых белков. Это хроматография на ДЭАЭ-целлюлозе, сефарозе или гидроксила-патите, гель-фильтрация, центрифугирование в градиенте плотности сахарозы и т. д. Очень важен правильный выбор детергента, поскольку именно детергент разрушает биомембрану, занимая место липидов, окружающих тот или иной белок, и определяет стабильность белка в растворе. Механизмы действия детергентов рассмотрены в обзоре.
2.1. ДЕТЕРГЕНТЫ
В течение последних двух десятилетий появилось очень много детергентов, пригодных для очистки интегральных мембранных белков. В принципе нужно пытаться найти такой детергент, который не нарушал бы вторичную и третичную структуры мембранных белков, а лишь замещал бы большинство или все мембранные липиды, контактирующие с гидрофобными участками белковой молекулы. Конечной целью солюбилизации является встраивание белка в детергентиую мицеллу; последующая стратегия очистки состоит в разделении таких белково-детергентных комплексов.
Первая проблема — это подбор оптимальных условий солюбили-зации изучаемого белка. Детергенты, денатурирующие белки, не подходят для решения такой деликатной задачи. С другой стороны, многие детергенты недостаточно эффективно разрушают мембраны и образуют белоксодержащие смешанные мицеллы. Такие детергенты могут быть либо слишком гидрофобными, либо слишком гидрофильными для эффективного смешивания с мембранными липидами и — при достаточно высокой их концентрации — для превращения бислоя в глобулярные смешанные мицеллы. Сначала надеялись, что выбор необходимого детергента удастся систематизировать с помощью одного параметра, называемого гидро-фильно-липофильным балансом. Этот параметр, изменяющийся от 1 до 20, используется при получении сурфактантов в качестве меры относительной гидрофобности. Действительно, получены некие корреляции, из которых следует, что значение ГЛБ детергента может использоваться для предсказания его поведения в биологических системах. Вообще говоря, можно сказать, что детергенты со значением ГЛБ в диапазоне от 12,5 до 14,5 являются наиболее эффективными растворителями интегральных мембранных белков. Однако впоследствии выяснилось, что поиск оптимальных детергентов для определенного мембранного белка требует учета многих факторов и всегда должен сопровождаться эмпирической проверкой. Необходимо учитывать следующее.
1.Максимальная солюбилизация исследуемого белка. Критерием является переход белка в супернатант после центрифугирования, при котором происходит осаждение мембраны.
2.Солюбилизация белка в нужной форме. Обычно речь идет о сохранении его ферментативной активности, но иногда используются определенные спектральные характеристики или наличие конкретных белковых ассоциатов. Кроме того, необходимым условием является стабильность белка после солюбилизации. В некоторых случаях для поддержания биохимической активности вместе с детергентом добавляют экзогенные фосфолипиды. В качестве примера можно привести получение лактозопермеазы Е. coliи белка натриевого канала. Иногда для стабилизации белка после солюбилизации добавляют глицерол или другой полиол. Имеет смысл использовать также ингибиторы протеаз и проводить солюбилизацию в условиях, сводящих к минимуму вероятность их протеолитического расщепления.
3.Возможность использования детергента в данной методике. Необходимо прежде всего учитывать заряд детергента, поведение при данном значении рН, ККМ и размер мицелл детергента. Последние свойства особенно важны. Детергенты с низкой ККМ, образующие крупные мицеллы, не удаляются при диализе или ультрафильтрации из-за слишком низкой концентрации мономеров детергента. С практической точки зрения это означает, что если концентрировать белок с помощью ультрафильтрации, то будет возрастать и концентрация детергента с низкой ККМ, а это может привести к денатурации белка. По этой причине многие исследователи предпочитают использовать детергенты с высокими ККМ, например октилглюкозид, соли желчных кислот или более современные цвиттерионные детергенты. Весьма ценными являются полистиреновые смолы, такие, как биобидз SM-2. Они избирательно связываются с детергентами типа тритон Х-100, удаляют их из раствора и позволяют обойтись вообще без диализа. Еще один фактор, который необходимо учитывать, — это поглощение света детергентом. Некоторые детергенты, например тритон Х-100, поглощают в ближней УФ-области, что делает невозможным определение концентрации белка по измерению оптической плотности при длине волны 280 нм.
С учетом всех этих факторов становится понятно, почему во многих случаях при выделении интегральных мембранных белков приходится использовать разные детергенты. Например, для солюбилизации можно применять тритон Х-100, а разделение с помощью ДЭАЭ-целлюлозы лучше проводить в присутствии октилглюкозида. Детергенты можно менять на стадии хроматографии, во время центрифугирования в градиенте плотности, а в некоторых случаях — с помощью диализа. Следует иметь в виду, что детергент, непригодный для солюбилизации определенного белка, может быть очень эффективным для сохранения белка в растворе после замены детергента. Очистку почти всегда следует проводить при избытке детергента в растворе, в противном случае равновесие будет сдвинуто в сторону агрегации мембранных белков, а не в сторону образования белково-детергентных комплексов. В некоторых случаях подобная агрегация может быть даже желательна, и последняя стадия очистки может состоять в удалении детергента. Но, как правило, при недостатке детергента происходят необратимое осаждение и потеря белка.
Необходимость поддержания концентрации детергента на определенном уровне создает дополнительнее трудности помимо тех, с которыми обычно сталкиваются при очистке белков; о некоторых из них мы уже говорили. Проблемы возникают и при использовании стандартного метода высаливания при высокой концентрации сульфата аммония: во многих случаях белок осаждается в комплексе с детергентом и липидом. Поскольку солевой раствор имеет высокую плотность, а детергент в агрегате — относительно низкую, то при центрифугировании преципитат будет оставаться на поверхности. Важно помнить, что очистке подвергаются белково-детергентные комплексы, нередко со значительным количеством связанного фосфолипида. Это сказывается на качестве разделения при хроматографировании, а также на результатах характеристики конечного прорастворимых белков, нужно определить число и молекулярную массу полипептидных субъединиц, их стехиометрию, размер и, возможно, форму молекулы, а также, если это необходимо, биохимическую активность.
|
Детергент 1 В табл. 1 и 2 перечислены наиболее широко используемые детергенты и указаны их свойства, важные для обсуждаемых нами вопросов. Эмпирически наиболее эффективными являются: 1) неионные детергенты (тритон Х-100, октилглюкозид); 2) соли желчных кислот (холат, дезоксихолат); 3) цвиттерионные детергенты (CHAPS, цвнттергент). Но выбор детергента, наиболее приемлемого для солюбилизации и очистки определенного мембранного фермента, по-прежнему осуществляется методом проб и ошибок. | ККМ, мМ | Мол.масса | 1 Размер А мнцеллы | .грегационис число | х Удельный объем, мл/г | Ссылки |
| Долецилсульфат | 1,33 | 288 | 24 500 | 85 | 0,864 | |
| натрия | ||||||
| Холат натрия " | 3 | 408 | 2100 | 5 | 0,778 | (612, 1383] |
| Дезоксихолат | 0,91 | 392 | 23 000 | 55 | 0,771 | |
| натрия " | ||||||
| 0,11 | 538 | 68 000 | 12 | 0,973 | ||
| Тритои Х-100 2) | 0,24 | 628 | 90 000 | 140 | 0,908 | |
| Твин 80 2) | 0,012 | 1300 | 76 000 | 60 | 0,8% | |
| Лаурилдиметил- | 2,2 | 229 | 17 000 | 75 | 1,112 | |612] |
| аминоксид | ||||||
| ^-D-Октил- | 25 | 293 | 8000 | 27 | 0,820 | [1242, 1213 |
| глюкозид | 612] | |||||
| ^-D-Лаурил- | 0,16 | 510 | 50 000 | 98 | 0,820 | |
| мальтозид | ||||||
| CHAPS | 8 | 615 | 6150 | 10 | 0,802 | |
| Цвиттергеит | 3,6 | 335 | — | — | 0,957 |
3. ХАРАКТЕРИСТИКА ОЧИЩЕННЫХ ИНТЕГРАЛЬНЫХ МЕМБРАННЫХ БЕЛКОВ
Характеристика очищенных мембранных белков, даже самых простых, может составлять определенные трудности. Как и в случае
3.1 МОЛЕКУЛЯРНАЯ МАССА СУБЪЕДИНИЦ
Электрофорез в полиакриламидном геле в присутствии додецил-сульфата натрия — это обычная методика, но в случае интегральных мембранных белков при ее применении возникают особые проблемы. В этом методе додецилсульфат связывается с полипептидными цепями, и комплексы белок—ДНС разделяются в полиакриламидном геле в соответствии с их стоксовыми радиусами, которые в большинстве случаев зависят от молекулярной массы. Молекулярную массу определяют, сравнивая электрофоретическую подвижность данного комплекса и известного стандарта. Однако связывание ДСН с неизвестным белком может качественно отличаться от связывания со стандартами, и тогда будет получен неправильный результат. Подобная ситуация наблюдается для интегральных мембранных белков с высоким содержанием неполярных аминокислотных остатков. С большинством растворимых белков ДСН образует комплексы в соотношении 1,4 г ДСН на 1 г белка, а с белками, содержащими большой процент неполярных остатков, может связываться больше детергента. Возникающий при этом дополнительный отрицательный заряд приводит к аномальному повышению электрофоретической подвижности, и определяемая молекулярная масса оказывается меньше, чем на самом деле. Возможна и другая ситуация. Связывающийся с ДСН мембранный белок может находиться в не полностью развернутом состоянии, что тоже приведет к аномальному повышению электрофоретической подвижности из-за образования более компактного комплекса белок—ДСН. Все эти эффекты весьма существенны. Например, лактозопермеаза имеет кажущуюся мол. массу 33 ООО, если измерять ее с помощью электрофореза в ПААГ в присутствии ДСН; в действительности же, как показывают результаты генетического анализа, ее мол. масса равна 46 ООО. Во многих случаях удается оценить молекулярную массу более точно, если построить график Фергюсона, представляющий собой зависимость электрофоретической подвижности от содержания акриламида как для стандартных белков, так и для исследуемого белка. Этот график зависит от радиуса Стокса и в меньшей степени — от заряда комплекса. Например, по результатам электрофореза в 12%-ном акриламидном геле одна из субъединиц цитохромно-г о комплекса Е. coliимеет кажущуюся мол. массу 28 ООО, а из графика Фергюсона получается величина 43 ООО, что совпадает с мол. массой, рассчитанной по данным о секвенировании соответствующей ДНК.
Еще одна проблема — возможное наличие четвертичной структуры. Некоторые мембранные белки агрегируют даже в присутствии ДСН. Например, гликофорин А или белок оболочки бактериофага М13 при электрофорезе в полиакриламидных гелях с ДСН находятся в основном в виде димеров. Иногда агрегация еще более усиливается при нагревании смеси белок—ДСН. Такая картина наблюдается, например, для субъединиц как митохондри-альной, так и бактериальной терминальных оксидаз. Чтобы оценить способность белка к необратимой агрегации, следует провести сравнительный анализ результатов электрофореза в полиакрила-мидном геле с ДСН для прогретых и непрогретых проб. Сходная проблема иногда возникает из-за присутствия детергента, использованного при очистке мембранного белка. Этот детергент необходимо удалить и заменить на ДСН, поскольку в некоторых случаях наблюдается четкая зависимость электрофоретической подвижности от присутствия детергента, с помощью которого солюбилизнровали фермент.
Итак, есть основания думать, что оценка молекулярной массы субъединиц сильно неполярных интегральных мембранных белков, определенная с помощью электрофореза в ПААГ с ДСН, может оказаться неверной. К несчастью, простая альтернатива этому методу отсутствует, и правильную величину часто получают либо по данным о полной первичной последовательности, либо с помощью точного гидродинамического анализа.
3.2 ОПРЕДЕЛЕНИЕ МОЛЕКУЛЯРНОЙ МАССЫ НАТИВНОГО БЕЛКА С ПОМОЩЬЮ ГИДРОДИНАМИЧЕСКИХ МЕТОДОВ
Применение этих методов для мембранных белков может быть сопряжено с большими трудностями, вызванными связыванием детергента. Чтобы оценить это в полной мере, рассмотрим вначале простой растворимый белок, для которого установлена мол. масса субъединиц с помощью электрофореза в ПААГ с ДСН и необходимо узнать, чем он является в неденатурированной, активной форме — мономером, димером или олигомером более высокого порядка. Для определения молекулярной массы белков часто используется гель-фильтрация, включающая сравнение со стандартными белками; здесь возникают проблемы, связанные с тем, что все стандартные белки имеют глобулярную форму, а исследуемый белок может быть не глобулярным, а слегка удлиненным. Такой белок с мол. массой 50 000 может элюировать со скоростью, соответствующей мол. мае
се 100 ООО. В связи с этим колонка для гель-фильтрации должна быть прокалибрована в соответствии со значениями радиуса Стокса, т. е. с размерами «эквивалентной гидродинамической сферы», а кроме того, параллельно необходимо использовать какой-либо другой метод. Обычно измеряют скорость седиментации с помощью либо аналитического ультрацентрнфугирования, либо центрифугирования в градиенте плотности сахарозы. Коэффициент седиментации равен
![]()
где м — молекулярная масса белка,
v — его парциальный удельный объем, ij — вязкость раствора, б — плотность раствора.
--> ЧИТАТЬ ПОЛНОСТЬЮ <--