Реферат: Метамерия или сегментация в живой природе

Opabinia - существо неясного систематического положения
В направлении объединения сегментов тела в более крупные отделы – тагмы, трилобиты пошли несколько дальше. Максимальное число сегментов туловища у трилобитов – 42. У вышестоящих форм начинается дифференцировка туловища на две тагмы: грудь (торакс) и пигидий. Происходит этот процесс путём слияния тергитов задних сегментов в один пегидиальный щит, тогда как сегменты груди остаются свободными и подвижными. У многих трилобитов на поверхности пигидия ещё остаются поперечные борозды, указывающие на его метамерное строение, но у других они полностью исчезают, и пигидий представляется с дорсальной стороны совершенно однородным.
Вторую большую группу членистоногих представляют CHELICERATA . Для подтипа хелицеровых характерно слияние головных и грудных сегментов, в результате чего образуется единая головогрудь, состоящая из семи слившихся сегментов. Головогрудь несёт шесть пар конечностей, первая из которых видоизменилась в хелицеры, вторая пара конечностей видоизменилась в педипальпы. Остальные четыре пары являются ходильными ногами.
Их иногда разделяют на три класса: EURYPTEROIDEA , XIPHOOOSURA, ARACHNOIDEA. Все хелицеровые обладают высоко гетерономной
 |  |
метамерией.

Ордовикский и силурийский ракоскорпионы, самые опасные морские хищники тех времен. Возможно, первые рыбы были "панцирными" для защиты от ракоскорпионов
А это уже настоящий скорпион, тоже силурийский. Очень похож на современных сухопутных скорпионов, хотя жил в воде. Это еще один пример необычайно долгого эволюционного стазиса.
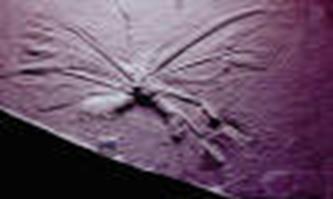
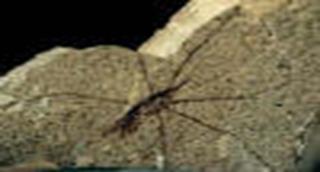
Эвритепиды существовали в течении большей части палеозоя. Имели продолговатое тело, на котором можно различить просому (передний отдел), покрытую головным щитом, и оптисому (задний отдел). В просоме, если рассматривать животное с брюшной стороны, можно найти, как и у трилобитов, верхнюю губу, сидящую впереди рта. Позади рта и позади последней пары ног располагается метасом, вероятно образованный слиянием рудиментарных конечностей седьмого сегмента. Кроме того, просома несёт шесть пар конечностей. Из них первая пара – хелицеры, обычно трёхчленистые – заканчивалась клешнями и служила для хватания. Остальные конечности – локомоторные и жевательные. Оптистосома распадается на два отдела, заметные при её рассмотрении с брюшной стороны. Первый отдел носит название мезосомы. Их первая пара представлена половыми крышечками. Следующие сегменты мезосомы (3-6) несут жаберные ножки. Шесть последних сегментов образуют метасому.
Просома мечехвостов покрыта большим щитом. Она несет шесть пар конечностей, причём пять из них снабжены клешнями и лишены эпиподитов, тогда как шестая заканчивается несколькими шипами и на своей прекоксе несёт нечленистый преэпиподит. У палеозойских форм все ноги были без клешней. Позади шестой пары имеется ещё одна пара нерасчленённых придатков – хиларий. Они являются рудиментами конечностей седьмого сегмента и развиваются за счёт их жевательных отростков. Рот сильно сдвинут назад, что нарушает гомономность метамерии. Хелицеры и педипальпы двигаются впереди рта. Четвёртый сегмент вытянут почти поперёк тела, пятый и шестой изогнуты в виде дуг, открытых спереди. Юрский и современный мечехвосты - почти одинаковые Большая медиальная часть сегмента входит в состав просомы, тогда как его плевры частично
 |  |
оказываются в составе мезосомы.
Седьмой сегмент рудиментарен, состоит из глабеллярного участка и в основном вошел в состав просомы, тогда как плевры его либо редуцированы, либо слились с задней частью плевры шестого сегмента. Конечности первого сегмента представляют собой половые крышечки, сегмент несёт половое отверстие, все остальные сегменты несут жаберные ножки. Заканчивается тело мечехвоста громадным шипом, подвижно сочленённым с последним сегментом мезосомы, метасома не выражена.
Древнейшим скорпионом является силурский водный скорпион PALAEPHONUS . Во многих отношениях он сходен с эвриптеридами. Просома несёт шесть пар конечностей: короткие клешневидные хелицеры, длинные клешневидные педипальпы и четыре пары ходильных ног. По расчленению тела скорпионы ничем принципиально не отличаются от эвритепид и близок к ним по набору конечностей. Анатомическая близость настолько велика, что скорпионов можно назвать наземными эвриптеидами.
Наземные хелицеровые, или паукообразные, отличаются значительной древностью: многие отряды паукообразных известны уже с карбона.
 |
Паук с мухой в янтаре
 |
Avicularia avicularia ( Птицеяд обычный ). Самка – до 50 мм, самец – до 35 мм.
Общее число сегментов в различных отрядах может различаться. Своеобразную черту в метамерию всех хелицеровых вносит судьба седьмого сегмента просомы. Только у LIMULUS он несёт рудиментарные конечности. В других отрядах он выражен в весьма значительной степени. Конечности просомы остаются ротовыми и локомоторными, конечности мезосомы – дыхательными.
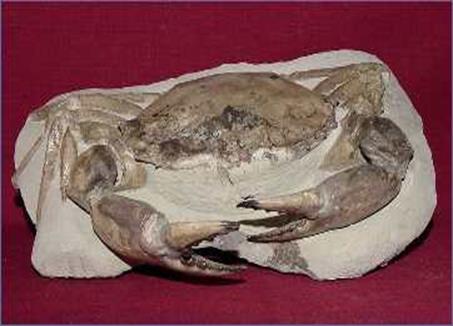 |
Внешнее строение ракообразных может быть весьма разнообразным. Тело у них расчленено на три отдела: голову ( cephalon ), грудь ( thorax ), и брюшко ( abdomen ). Строение головы может иметь свои особенности. На сегментах головы могут располагаться различные придатки, которые могут быть парными (антенны и видоизменённые конечности) и непарными (верхняя губа). Сегменты головы несут конечности, которые гомологичны параподиям полихет. Остальные три сегмента головы также несут конечности, но они обычно видоизменяются в ротовые части и выполняет функции первичной переработки пищи. Конечности второго сегмента преобразуются в ротовые челюсти. Конечности грудных сегментов также специализируются. У многих
Миоценовый краб
видов конечности первых трёх пар грудных сегментов преобразуются в ногочелюсти первых
 |
трёх пар грудных сегментов. Остальные ноги служат для передвижения. Брюшные сегменты несут конечности только у высших раков, при
Миоценовый краб
этом они специализируются не для
передвижения, а для дыхания или размножения
и у самцов и самок могут иметь различное
строение. Последняя пара брюшных конечностей (уроподы) видоизменена в уплощенные пластинки, которые вместе с тельсоном образуют широкий плавник.
Среди ракообразных наиболее примитивный план строения, равно, как и гомономную сегментацию сохранили ANOSTRACA , которые являются примитивнейшими из членистоногих вообще, но туловище ракообразных никогда не достигает той степени гомономности, которая присутствует у трилобитов.
 |
Юрская креветка с очень длинными антеннами
У высших ракообразных голова состоит первичной ( PROTOCEPHALON ) и вторичной ( GNATHOCEPHALON ) головы. Максиллярная область головы очень часто образует щит или двустворчатую раковину в виде складки, покрывающей переднюю часть туловища. Она служит не только для защиты, но и является струенаправляющим органом. Позади них хорошо заметен тергит мандибулярного сегмента, тогда как максиллярные тергиты, за счёт которых образуется и свободная часть щита, полностью слиты между собой. Туловище распадается на два отдела: грудь и брюшко. У большинства грудь состоит из тринадцати сегментов, из которых одиннадцать сегментов несут по одной паре одинаковых листовидных ножек, тогда как конечности двенадцатого и тринадцатого сегментов превращены в половые придатки, тринадцатый сегмент несёт половое отверстие. Брюшко состоит из шести безногих сегментов, за которыми следует анальная лопасть – тельсон, несущая парные придатки, образующие вилку. Грудные ноги нерасчленённые, листовидные, на наружном крае несут один или два преэпиподиита, один эпиодит и нерасчленённый экзоподиит. На внутреннем крае они несут жевательные эндиты. Чрезвычайно разнообразна метамерия туловища у NOTOSTRACA . Если первые одиннадцать сегментов груди имеют по одной паре ножек, то последующие 16-20 сегментов несут по несколько пар – от двух до шести на каждом сегменте.
 |
Третий большой ствол членистоногих образуют ATELOCERATA (насекомые и многоножки). Они являются самыми
Kalligramma (отр. сетчатокрылые - златоглазки и муравьиные львы; ранняя пермь - ныне)
многочисленными из животных вообще. Тело насекомых чётко разделено на голову, брюшко и грудь, подвижно соединённые между собой. Голова состоит из акрона и четырёх слившихся сегментов, окружённых общей хитиновой капсулой. Придатками акрона является пара антенн, гомологичных антенуллам ракообразных. Первый сегмент головы редуцирован, в результате чего вторая пара антенн не образуется. Придатки остальных трёх сегментов головы являются видоизменёнными конечностями, которые в своей совокупности образуют ротовой аппарат. В его состав входят: придатки второго сегмента – мандибулы, придатки третьего сегмента – первая пара нижних челюстей. Сросшиеся придатки четвёртого сегмента – вторая пара нижних челюстей, которые вместе образуют вторую непарную губу. Грудь у всех насекомых состоит из трёх сегментов, которые называются переднегрудью, среднегрудью, и, соответственно, заднегрудью. Все они несут по одной паре ходильных ног, а последние два сегмента несут ещё и по паре крыльев. Брюшко может содержать разное количество сегментов, причём у примитивных форм сегментов всегда больше. Конечности на брюшке отсутствуют, но у примитивных форм, а также у тараканов они сохраняются в виде придатков. Наиболее примитивные обладают сегментацией, близкой к гомономной. Тело всех многоножек разбивается только на два отдела: голову и туловище, которое очень часто состоит из очень большого числа сегментов, из которых все, или почти все несут по паре более или менее одинаковых конечностей.
Chresmoda (водомеркоподобное насекомое неясного положения)
 |
Стрекоза Cymathophlebia
 |
Голова всех многоножек представляет результат слияния акрона и четырёх сегментов. Она представляет собой монолитное образование без следов метамерного происхождение, если не считать наличия нескольких пар конечностей. У насекомых всегда сохраняются интерсегментальные швы, в том числе и у многоножек. Редукция конечностей первого постантеннулярного сегмента – признак, общий для всех ATELOCERATA . Этому признаку они обязаны своим названием. Второй постантеннулярный сегмент несёт хорошо развитые конечности, представляющие собой жвалы, или мандибуллы. Конечности третьего и четвёртого сегментов представлены двумя парами максилл. Голова насекомых по своему составу не отличается от головы SYMPHYLA , а также несёт конечности трёх челюстных сегментов. Характерной особенностью насекомых является разбивка их туловища на два отдела: грудь и брюшко. Грудь берёт на себя локомоторную функцию: здесь сохраняются ноги и у высших насекомых возникают крылья. Брюшко, наоборот, теряет все конечности, если сохраняются, то только в виде половых придатков. Грудь почти всецело берёт на себя двигательную функцию. В силу этого мускулатура грудных сегментов увеличивается, наружный скелет осложняется. В сегментах брюшка наоборот. У хорошо летающих насекомых происходит всё возрастающая консолидация груди, интеграция её сегментов в одно сложное целое. Таким образом от почти гомономного туловища наиболее примитивных насекомых, мы видим ряд переходов к гетерономному и интегрированному туловищу.
Гомология (греч. homologia — соответствие) сходство органов, построенных по одному плану и развивающихся из одинаковых зачатков у разных животных и растений; такие гомологичные органы могут быть неодинаковы по внешнему виду и выполнять различные функции. Определение гомологии и противопоставление её аналогии предложил английский учёный Р. Оуэн (1843), различавший частную гомологию, то есть соответствие органа по положению и связи с другими частями органу другого животного (например, рука человека, ласт кита, крыло птицы и др.), и сериальную гомологию, или гомодинамию, то есть соответствие у одного и того же животного частей тела, расположенных по длинной оси (например, рука и нога человека). Естественноисторическое объяснение гомологии общностью происхождения организмов впервые дал Ч. Дарвин (1859). Немецкий анатом К. Гегенбауэр (1898) различал полную гомологию — сходство органов по их положению и связям с другими органами не нарушается вариациями в форме и величине: и неполную гомологию, при которой отдельные части органов могут в процессе эволюции исчезать (дефектная гомология) или появляются новые части (аугментативная гомология). Комбинацию утраты одних и новообразования других частей организма назвали имитативной гомологии (немецкий биолог М. Фюрбрингер). Морфологический критерий гомологии — одинаковые положение и строение органов, а также наличие между ними переходных форм. Онтогенетический критерий гомологии — развитие органов из сходных зачатков. Пример гомологии органов у растений: видоизменённые в связи с выполнением разных функций листья, превратившиеся в лепестки цветка, тычинки, некоторые виды колючек. Частные случаи гомологии — гомодинамия , гомономия , гомотипия . Гомологичные органы животных имеют одно и то же эволюционное происхождение независимо от выполняемой у данного вида функции. Это, например, руки человека и крылья птиц или хвосты рыб и обезьян, которые одинаковы по происхождению, но используются по-разному.
Аналогичные структуры сходны по выполняемым функциям, но имеют разное эволюционное происхождение. Это, например, крылья насекомых и птиц или ноги пауков и лошадей. Органы могут быть гомологичными и