Реферат: Метамерия или сегментация в живой природе
Около 200 лет назад великий французский естествоиспытатель Ж. Кювье, работая над созданием системы животного мира, выделил шесть типов животных, в т. ч. тип членистых, в который он объединил всех существ, тело которых разделено на сегменты: насекомых, раков, пауков, мокриц, дождевых червей и пиявок.
В ходе эволюционного развития появлялись виды плоских червей, которые увеличивали длину своего тела, отращивая сегмент за сегментом. Эти существа просто добавляли очередное мускульное колечко. Но затем изменения охватили все внутреннее строение червей. Их органы также стали расти частями. В результате появились животные с наружным и внутренним членением тела, в котором почти каждый новый сегмент в общих чертах копирует предыдущий. Такое повторение однородных сегментов тела получило в науке название метамерии.
В действительности, конечно же, развитие метамерного строения тела протекало гораздо дольше и сложнее. Еще у примитивных микроскопических корненожек фораминифер наблюдается подразделение раковинок на сходственные сегменты. Таким образом, сегментация тела лежала в основе эволюции органического мира. Она была следствием приспособления живых существ к среде обитания и эволюции типов симметрии тела, увеличивающей адаптационные возможности
 |
Нитевидная форма Palaeolyngbya из позднего протерозоя Австралии
(850 млн. лет).
видов. Сама по себе метамерия рассматривается учеными как особый тип симметрии.
 |
Медузы и их сородичи из типа кишечнополостные, а также низшие черви лишены настоящих метамеров (сомитов). Однако многим им свойственны либо полимерные органы, либо псевдометамерия, то есть, наличие ложных сегментов. У таких
 |
Девонская колониальная ругоза - Hexagonaria
Кайнозойский шестилучевой коралл Meandrina
кишечнополостных, как гидрозои, сци-фозои, антозои и ктенофоры, имеются полимерные органы.
Обычно полимерами называются химические вещества, родственные полиэтилену. В биологии такое же наименование получили органы и структуры, неоднократно повторяющиеся в организме. Обычно это органы половой системы. Псевдометамерия и полимерные образования типичны для ряда групп низших червей: цестод, турбеллярий, немертин. У сколецид закладываются полости тела, отчего и возникают подобия сегментов.
Дальнейшее разделение тела животных оказалось возможным благодаря усложнению нервной системы и, главным образом, появлению кровеносной системы. Стало возможным обособление разветвлений нервов (с формированием невросомитов) и кровеносных сосудов (с формированием ангиосомитов).
Третьим и, пожалуй, наиболее существенным условием прогресса метамерии стал рост мускулатуры. Кольчецы обладают наиболее развитым среди червей мышечным мешком, объем которого достигает 60-70% от общего объема тела. За счет интенсивного развития мышц произошло обособление участков продольной и кольцевой мускулатуры с формированием миосомитов, разделенных поперечными перегородками-диссепиментами.
Рост мускульного мешка и сопутствующая этому явлению сегментация были единым эволюционным рывком кольчецов, позволившим им занять важнейшие экологические ниши моря, пресных вод и почвы из тех, которые только занимают черви. Сколециды на протяжении всей своей истории боролись за надежную защиту и скорость. Лишь потомки примитивных червей смогли решить эту задачу, чем и объясняется прогресс этих групп. Улитки "изобрели" для себя надежную защиту в виде раковин. Кальмары создали из своего мускульного мешка мощный водометный "двигатель".
Кольчатые черви обрели плотные покровы, насеченные на дольки - сегменты, соответствующие внутренней метамерии. Сегментация позволила этим беспозвоночным свободно изгибать тело, чтобы передвигаться волнообразно, развивая при этом значительную скорость. Впоследствии насекомые, раки и прочие членистоногие покрылись
плотным сегментированным панцирем.
Метамерию можно рассматривать как частный случай симметрии. Метамерносимметричным является такое тело, которое совпадает само с собой при передвигании его вдоль прямой линии – оси переноса, или оси метамерии – на некоторое расстояние, обозначаемое, как длина метамеры. Метамерию можно обозначить, как поступательную симметрию.
Кроме обычной метамерии, вызванной наличием простой оси переносов, в органическом мире встречаются ещё два более редких типа метамерии: один с наличием винтовой оси переноса, другой – сочетание поступательной симметрии, наличием плоскости скользящего отражения.
Винтовая ось распространена как элемент симметрии стебля у сосудистых растений. При этом угол вращения на один шаг переноса (на одну метамеру) определяет собой порядок винтовой симметрии. Среди METAZOA винтовую симметрию в расположении частей обнаруживают, например, головки гидроида STAUROCORUNE FILIFORMIS . Вытянутая головка гидранта несёт 7-9 метамерно расположенных колец головчатых щупалец. В каждом кольце по 4 щупальца. Кольца чередуются так, что каждое вышележащие повёрнуто на 45 градусов по сравнению с соседним.
При наличии плоскости скользящего отражения каждая последующая метамера представляет зеркальное отражение предыдущей, и, таким образом вдоль всего тела животного чередуются «правые» и «левые» метамеры. Отдельные метамеры при этом типе метамерии сами по себе диссиметричны. В наиболее чистом виде скользящая метамерия встречается в стробиле цестоды TATRIA MATHEVOSSIANAE . Появление этого типа метамерии обнаруживается в строении мускулатуры и отхождении спинномозговых нервов ланцетника и в некоторых других случаях.
Метамерия, как особый вид симметрии могла бы существовать лишь при том условии, если бы ряд метамер в обе стороны тянулся бы до бесконечности. Так как ряд метамер всегда является конечным, то при передвигании его вперёд его передняя метамера не совпадает ни с чем. Поэтому метамерия животных и растений, если рассматривать её как форму симметрии, всегда является несовершенной.
Метамерия может быть полной и неполной . Метамерия является полной в том случае, если повторение охватывает всю организацию животного, и неполной, если она охватывает одни аппараты, одни сегменты органов и не охватывает другие.
Метамерия может быть гомономной и гетерономной . Метамерией в полном смысле слова является лишь гомономня метамерия. Гетерономная метамерия возникает в том случае, если метамерные образования дифференцированы, и таким образом перестают быть вполне сходными между собой, хотя и сохраняют общий план строения.
Метамерия может быть простой и сложной . О сложной метамерии говорят в тех случаях, когда составляющие тело животного метамеры располагаются повторяющимися группами, построенными по одному плану, так что эти группы являются как бы метамерами 2-го порядка.
Метамерия может возникать в организации животных несколькими путями: метамерным упорядочением первоначально хаотично расположенных гомотипных частей, (принадлежащих к одному и тому же типу частей); метаменрной дифференцировкой вдоль первоначально однородного целого; соединение первоначально независимых гомотипных частей . Первый и второй способ возникновения метамерии имеют между собой то общее, что здесь метамерное целое возникает из такого же целого либо вовсе лишенного метамерии, либо обладающего неопределённой поступательной симметрией. В третьем способе метамерное целое возникает из совокупности независимых между собой неметамерных единиц низшего порядка.
Метамерию мы встречаем во всём животном царстве. У простейших она наиболее ярко выражена у FOR А MINIFERA . Здесь она возникает путём неполного деления и выражается в строении раковины, состоящей из ряда метамерно расположенных камер. Эта метамерия является несколько гетерономной, так как камеры не имеют одинакового размера. У CILIATA метамерия развивается путём стробиляции – возникновением метамерного целого путём незавершённого до конца поперечного деления. Деление особи происходит очень быстро и не успевает дойти до конца, как наступает следующее деление. Возникшие цепочки непрерывно распадаются, но животное всё равно сохраняет вид цепочки. Существует также метамерия колоний . У турбеллярий метамерия иногда возникает путём упорядочения – благодаря тому, что кишечник имеет разветвлённое строение половые железы, лежащие в свободных промежутках, приобретают столь же правильное расположение. Нервный аппарат в силу этого приобретает такое же метамерное строение.
У кольчатых червей ( ANNELIDA ) существует два способа роста: олигомерное (все сегменты полностью закладываются на стадии личинки, их число не велико и их дельнейшего образования не бывает) и полимерное (на протяжении оральной стороны закладывается лишь часть сегментов. В результате этих процессов создаётся вторая личиночная стадия – метатрохофора или нектохета. Вслед за тем у личинки образуется зона роста, в которой идёт усиленное деление клеток, ведущее к образованию нового отдела туловища). У олигомеров метамерия не распространяется на целомическую мезодерму, в то время как у полимеров она бывает метамерной. Существует три теории происхождения метамерии у аннелид: теория происхождения метамерии аннелид от антимерии кишечнополостных, теория возникновения метамерии путём метамерной дифференцировки и упорядочения органов, теория происхождения её путём подавленного поперечного деления.
В то время когда для кольчатых червей характерна, в основном, гомономная метамерия и гетерономность в пределах этого типа играет лишь подчинённую роль, членистоногие являются животными гетерономно-метамерными. В тоже время и самый характер гетерономности несколько различен, что в значительной мере зависит от различий в характере покровов и функциях конечностей. Наличие толстой кутикулы у членистоногих ведёт к образованию щитов, объединяющих группы смежных сегментов в отделы и слиянию сегментов между собой. Параподии многощетинковых выполняют локомоторную, гидрокинетическую, дыхательную и чувствительную функции. Примитивные конечности членистоногих несут, кроме того, хватательною и жевательную функции. Для членистоногих характерно постоянное использование конечностей для хватания и жевания. Примитивность метамерии трилобитов заключается, прежде всего, в чрезвычайной гомономности их конечностей. За исключением нитевидных антенн остальные конечности у трилобитов по одному общему типу. Они сравнительно мало различаются у разных видов и очень сходны на различных сегментах тела. Конечность трилобита состоит из предкоксы, коксы, шестичленного телоподита и многочленистого, несущего жаберные листочки, преэпиподита, прикрепляющегося к предкоксе. Кокса, а иногда и часть члеников телоподита вооружены жевательными и хватательными эндитами.
Marella - одна из самых
массовых форм в уникальном
 | 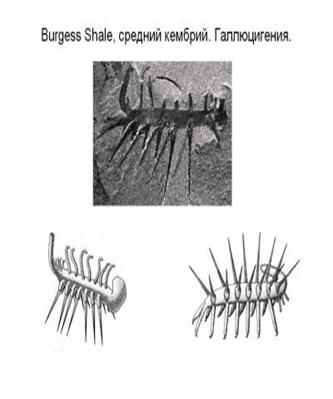 |
????????????????? ??????????????? Burgess Shale
Ротовой диск Anomalocaris
--> ЧИТАТЬ ПОЛНОСТЬЮ <--