Реферат: Молекулярные механизмы передачи импульса в мембранах нейронов. Ионные каналы, рецепторы
Общая схема молекулярной организации кальциевых каналов сходна с описанной выше для Na-каналов. Однако главная а-субъединица окружена большим числом субъединиц, служащих модуляторами активности канала. Пока не ясно, какие химические группировки ответственны за трансмембранный перенос кальция, понятно только, что он - существенно зависит от внутриклеточной концентрации Са+ и функционирования системы циклических нуклеотидов.
Несмотря на то, что численность кальциевых каналов значительно меньше, чем натриевых и калиевых ион-транспортных систем, при определенных условиях они могут самостоятельно вызывать деполяризацию нейрона. Однако сейчас очевидно, что главная функция кальциевых каналов состоит в сопряжении электровозбудимости с внутриклеточными процессами. Эта функция кальциевых каналов особенно важна для включения механизма выхода нейромедиатора из нервного окончания.
5. СИСТЕМЫ АКТИВНОГО ТРАНСПОРТА ИОНОВ. Na+ /K+ И Na+ /Ca+ - НАСОСЫ
Как уже упоминалось, электрическое возбуждение в нервной ткани существенно зависит от механизмов пассивного и активного мембранного транспорта, контролирующего концентрации ионов и молекул внутри клеток и нередко в межклеточном пространстве. Аксоны обладают большим резервом Кононов и дефицитом Na+ -noHOB. Миграции ионов, обеспечивающие прохождение импульсов и создающие изменения потенциала мембраны, быстро компенсируются этими резервами. "Выносливость" аксона очень велика — утомление наступает лишь после прохождения 10 — 10 импульсов, тем не менее перемещения ионов при прохождении импульса должны быть компенсированы в стадии покоя. Кроме того, мембрана в стадии покоя не является абсолютным барьером для перемещений ионов и постепенного уменьшения потенциала.
Ряд внутриклеточных процессов требует постоянной регуляции за счет активных ионных потоков через -мембрану. Наконец, особенно велики нарушения градиентов ионных концентраций при функционировании синапсов. Для компенсации всех этих нарушений градиентов служат ионные насосы.
Основной транспортной системой в нейронах, как и в большинстве других эукариотических клеток, является насос, который вытесняет Na+ и постоянно накапливает К+ . Этот процесс требует присутствия АТФ и специфически ингибируется кардиоактивными гликозидами типа оуабаина.
Na+ , К+ -активируемая аденозинтрифосфотяза, или АТФ-фосфогидролаза К.Ф.3.6Л плазматических мембран нервных клеток осуществляет трансмембранный перенос одновалентных катионов против градиентов их электрохимических потенциалов, используя энергию гидролиза АТФ. Работая с максимальной скоростью, этот ферментативный комплекс способен транспортировать через мембрану около 200 ионов Na и 130 ионов К в 1с. Однако фактическая скорость работы фермента определяется потребностями клетки. У большинства нейронов на поверхностной мембране расположено до 200 натриевых насосов на квадратный микрон, причем в некоторых участках этой поверхности их плотность в 10 раз выше.
Молекула фермента состоит из каталитической а-субъединицы и $-гликопротеида, функциональная роль которого до сих пор неизвестна. Молекулярная масса комплекса белковых субъединиц Na+ , К+ -АТФазы составляет около 275 000, и размеры фермента колеблются в пределах 6-8 нм. Кроме полипептвдов Na+ , К+ -АТФаза содержит 3 углеводные цепи, которые присоединены к р-субъединице гликозидными связями. Одновременное использование методов генной инженерии и химии белка привело к установлению первичной структуры Na+ , К+ -АТФазы. С помощью моноклональных антител установлено внутри- и внеклеточное расположение некоторых участков а- и р-субъединиц. Предложена модель полипептидной цепи фермента, согласно которой сс-субъединица 7 раз пересекает бислойную мембрану и локализована главным образом в цитоплазме. свидетельствует в пользу того, что существуют общие функциональные и структурные взаимодействия между Na+ , К+ -АТФазой и ионными каналами для транспорта К+ .
Сходным с Na+ , К+ -АТФазой является другой ионный насос — Ка+ /Са+ -АТФаза. Она производит обмен каждого иона Са+ на 3 иона Na+ . Значение этой системы особенно велико в нервных окончаниях, где система медиаторов связана с вхождением Са+ в терминаль и необходимостью компенсировать далее эти смещения градиента. Кроме того, ряд событий в постсинапти-ческой зоне тоже сопряжен с временным вхождением Са+ .
6. МОЛЕКУЛЯРНЫЕ ОСНОВЫ СИНАПТИЧЕСКОЙ ПЕРЕДАЧИ, РЕЦЕПТОРЫ
Нейрон способен иметь до нескольких десятков тысяч межклеточных контактов, большинство из которых обеспечивается определенными морфологическими структурами — синапсами. Клеточную поверхность нейронов можно рассматривать как приемник разнообразнейших сигналов.
Чем выше степень эволюционной организации нервной системы, тем разнообразнее природа химических синапсов. Особенно это касается головного мозга высших млекопитающих, включая человека. Очевидно, химические синапсы оказались эволюционно более выгодными для передачи дискретных сигналов по сравнению с другими типами межклеточных контактов, поскольку на их 'основе возможна не только передача сигнала, ко и его разнообразная модуляция, в том числе гуморальными факторами. Основой восприятия нейроном химического сигнала в синапсе, а также ряда модулирующих влияний являются рецепторы.
Рецепторы представляют собой надмолекулярные образования, состоящие из белков, а также гликолипидных компонентов. Они способны под действием медиатора либо нашэсредственно изменять потоки ионов через мембрану, либо индуцировать образование вторичш>тх мессенджеров, которые, в свою очередь, меняют ряд свойста лейрона.
Межнейрональные химические синапсы подразделяются на два типа: возбуждающие и тормозные, причем.первые, как известно, способствуют генерации новых импульсов, а вторые приводят к снятию действия приходящих сигналов. Это деление определяется в значительной мере природой рецепторов. Известны случаи, когда один и тот же медиатор оказывает возбуждающее или тормозное действие в зависимости от природы рецептора.
В зависимости от места положения синапсов их можно подразделить на сомато-аксональные, дентрито-аксональные, ден-трит-дентритные и др. Каждый из этих синапсов имеет свои особенности в функционировании. Схематически структура синапса может быть представлена следующим образом.
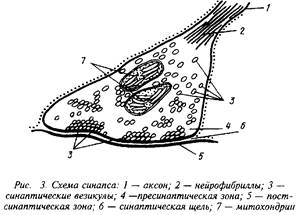
На рисунке хорошо видны утолщения, составляющие пре-синаптическую мембрану подходящего аксона, синаптическая щель и постсинаптическая мембрана. В пресинаптическом окончании находятся синаптические везикулы — хранилища запасов нейромедиатора в пресинаптическом нейроне. Постсинаптическая мембрана является носителем рецепторов. В ряде случаев сами рецепторы могут быть визуализированы при посредстве электронной микроскопии.
При распространении нервного импульса происходит деполяризация пресинаптической мембраны я изменение ионных токов. Наиболее важным событием в нервном окончании является мобилизация ионов Са, которые вызывают миграцию и открывание многочисленных синаптических везикул. Эти везикулы непосредственно связываются с участками пресинапса и открытие их приводит к высвобождению нейромедиатора и диффузии его в синаптическую щель. В терминали аксона сконцентрированы и ферменты синтеза медиатора, митохондрии для энергетического обеспечения этого процесса, системы белков-транспортеров, способствующих узнаванию и обратному захвату молекул нейромедиатора. Этот последний механизм, по-видимому, существенно экономит затраты на синтез готового нейромедиатора и участвует в регуляции срока его действия.
В отличие от биохимических процессов выброса нейроме-диаторов из пресинапса, имеющих общий характер, постсинап-тическое действие нейромедиатора прекращается самыми разнообразными способами: разрушением его определенными ферментами, либо быстрым поглощением из области синапса гли-альными клетками, либо обратным захватом его в пресинаптическую терминаль. В качестве примера существования разных механизмов утилизации нейромедиаторов из синапса можно привести механизмы процессов, происходящих в холинергиче-ском, глутаматергическом и ГАМК-ергическом синапсах.
Механизм разрушения ацетилхолина преимущественно связан с работой фермента — ацетилхолинэстеразы, который располагается на постсинаптической мембране и быстро гидроли-зует медиатор после взаимодействия с рецепторами. В глутаматергическом синапсе механизм удаления нейромедиатора заключается преимущественно в поглощении L-глутамата окружающими глиальными клетками. L-глутамат превращается в глутамин с помощью фермента глутаминазы, находящейся в глиальных клетках. В ГАМК-ергическом синапсе преобладает система обратного захвата медиатора.
7. ОБЩИЕ ВОПРОСЫ НЕЙРОРЕЦЕПТИИ
В биохимии и физиологии нервной системы длительное время доминировала точка зрения, согласно которой местом активных пластических изменений нейронов является пресинапти-ческая мембрана. Были представлены многочисленные свидетельства, касающиеся сдвигов количества квантов нейромедиатора в процессе обучения, памяти, выработки условных рефлексов и др. Постсинаптическим мембранам отводилась либо пассивная роль, либо они вообще не рассматривались в качестве активных участников событий в нервных клетках. Впоследствии стало ясно, что процессы, происходящие в плазматической или синаптической мембране нейрона, являются одними из ключевых для понимания интегративных функций ткани мозга, решения проблем обеспечения эффективного взаимодействия между нервными клетками. В последние десятилетия особое внимание было обращено на изучение структурных компонентов постсинаптических мембран, в частности нейрорецепторов. Исследования тонкой структуры и функции нейрорецепторов показали их важную роль в трансформации химического сигнала в биоэлектрические потенциалы и в передаче информации на внутриклеточные реакции, которые определяют метаболизм нервной ткани.
Следует отметить, что нейрорецепторы расположены как на мембранах нейронов, так и на мембранах глиальных клеток. Однако у последних они имеются в ограниченном наборе и числе. Рецепторные системы, расположенные на глиальных элементах, отличаются от нейрональных весьма важным моментом — они не способны продуцировать ответные реакции клеток в виде оперативных единиц информации. Иными словами, они не генерируют потенциалов действия. Как правило, глиальные клетки реализуют свое действие через внутриклеточные и межклеточные трофические регуляторные реакции, участвуя в метаболизме нейронглия.
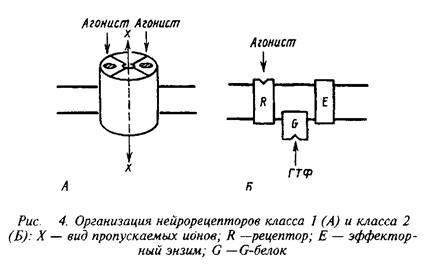
Несмотря на огромное разнообразие клеточных рецепторов на мембране нейрона, их можно подразделять на два основных класса, которые различаются по механизмам действия и скорости проведения сигналов. Существуют быстродействующие ионотропные и медленнодействующие метаботропные рецепторы скорость действия первых составляет миллисекунды, в то время как у последних они находятся в секундно-минутном диапазоне. Время действия нейрорецепторов определяется структурной организацией рецепторных компонентов.
Быстродействующие рецепторы содержат в своей структуре ионный канал, открывающийся при контакте с нейромедиатором. Медленнодействующие рецепторы представляют собой комплекс из нескольких белков, которые при воздействии нейромедиатора последовательно меняют конформацию и в конечном счете активируют синтез или выход вторичного, уже внутриклеточного, медиатора. Эти два класса рецепторов обозначают нередко как рецепторы I и II класса. Для правильного восприятия терминологии целесообразно также указать, не рассматривая пока детали, что рецепторы класса II содержат в числе белков, передающих сигнал, так называемые G-белки. Их нередко упоминают, обозначая рецепторы этого класса.
Кроме охарактеризованных выше двух классов рецепторов существуют еще три особые группы рецепторов, которые хотя и присутствуют в нервной системе, но пока представляются не связанными прямо со специфическими функциями последней. К ним относятся рецепторы, переносяшие свои лиганды через мембрану, рецепторы, обладающие собственной тирозинкиназной активностью и, наконец, своеобразная группа, которая при взаимодействии с лигандом претерпевает частичное протеолитическое расщепление. В настоящем руководстве мы не рассматриваем эти группы рецепторов.
К первому классу рецепторов принадлежат никотиновые рецепторы ацетилхолина, рецепторы ГАМКА , глицина, а также часть рецепторов глутамата и аспарагиновой кислоты. Рецепторы катехоламинов, серотонина, ГАМКВ и ряда пептидных соединений, а также мускариновые рецепторы ацетилхолина и некоторые из рецепторов глутамата относят ко второму классу. Последние типы рецепторов через систему вторичных посредников вызывают изменения в активности проте-инкиназ, способных фосфорилировать мембранные белки, включая ионные каналы.
Таблица 1