Реферат: Молекулярные механизмы передачи импульса в мембранах нейронов. Ионные каналы, рецепторы
Таблица 2
Масса белков рецепторов класса II
| Рецептор | Мг, кД |
| Ад рене р ги чески й | 58-80 |
| Глутаматер гически й | 90-110 |
| Холинергический | 85-105 |
| Дофаминовый | 72-94 |
| Опиатный | 53-65 |
| Серотониновьш | 67 |
Следует отметить, что в последние годы обнаружена группа нейрорецепторов, связь которых с ионными каналами осуществляется через G-белки, не сопряженные с перечисленными выше вторичными мессенджерами. Хотя в такую систему рецепции и не включены протеинкиназы, тем не менее участие G-белка в трансформации сигнала значительно увеличивает время действия по сравнению с нейрорецепторами класса 1.
Фундаментальным свойством всех нейрорецепторов является их лабильность и высокая скорость синтеза самого рецептора. Это свойство рецепторов конрастирует с более жесткой запрограммированностью синтеза белковых компонентов мембран, которая обычно наблюдается у других типов тканей. В нейронах развиты механизмы непрерывного синтеза рецепторов и их быстрой утилизации либо путем интернализации, либо с помощью пиноцитоза. Высокая скорость обновления нейрорецепторов обусловлена, по-видимому, необходимостью изменения "информационной емкости и "пропускной способности" нейрона. В этом случае генетический аппарат клетки способен, интегрируя всю приходящую информацию, "принять решение" путем перестройки синтеза белковых компонентов мембран. В этом скрыта одна из причин уникального свойства нейронов и нервной ткани в целом — пластичности.
Таким образом, основная роль нейрорецепторов сводится к созданию специфических информационных входов, организующих единый функциональный ансамбль нейронов. Именно совокупность рецепторов определяет лицо клетки и ее реакции на поступление разнообразных химических сигналов.
Молекулярные механизмы, лежащие в основе модуляции эффективности синаптической передачи, в которых важную роль играют рецепторные процессы, имеют альтернативу. С одной стороны, это изменение чувствительности к рецептору, с другой — увеличение или снижение количества активных рецепторов на мембране. Заслуживают внимания и гипотезы, касающиеся посттрансляционной модификации нейрорецепторов, которая позволяет изменить количественные параметры их функционирования.
Внимание к проблемам нейрорецептии со стороны биохимиков, фармакологов и физиологов обусловлено еще и тем, что причиной многих дисфункций нервной системы является нарушение целостности мембранных компонентов как нейронов, так и глиальных клеток. Отметим, что существующие успехи в лечении некоторых нервно-психических заболеваний связаны в большей мере с прогрессом в исследовании именно молекулярных свойств ряда рецепторов. Оказалось, что многие нейрорецепторы выполняют роль избирательных мишеней действия известных лекарственных препаратов. Исследования в этой области нейробиологии служат сейчас постоянным источником для целенаправленного поиска и создания новых классов фармакологических средств, обладающих улучшенными терапевтическими свойствами.
8. БИОХИМИЧЕСКИЕ МЕТОДЫ ИЗУЧЕНИЯ МЕХАНИЗМОВ НЕЙРОРЕЦЕПТИИ
Наиболее широко распространенным и разработанным методическим подходом для количественного анализа взаимодействия нейромедиаторов со своими рецепторами на мембране клетки является радиолигандный метод. Суть этого метода заключается в изучении параметров связывания радиоактивного лиганда с мембранно-связанными или изолированными рецепторными белками. В настоящее время существует хорошо развитая кинетическая теория рецептии и методы определения физико-химических параметров процесса образования комплекса лиганд-рецептор. Такой физико-химический анализ позволяет сделать определенные заключения о структуре активных центров нейрорецепторов, в частности, выяснить природу некоторых функциональных групп, которые ответственны за первую стадию взаимодействия лиганда с акцептором.
Для того чтобы кратко ознакомиться с количественной теорией взаимодействия веществ со своими рецепторами, рассмотрим простейшие условия, когда одна молекула лиганда взаимодействует с одним центром связывания:
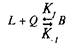
где L — лиганд; Q — центр связывания; В — комплекс лиганда со связывающим центром; К{ и K.j — кинетические константы. При динамическом равновесии скорость реакции образования комплекса В равна его скорости диссоциации, т.е. Vt = V_1? тогда концентрация вычисляется по формуле
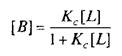
При этом предполагается, что L и Q взаимодействуют между собой по закону действующих масс, т.е. скорости реакций образования комплекса и его диссоциации прямо пропорциональны концентрациям компонентов в системе. Отношение констант прямой и обратной реакции называют константой сродства Кс . Она характеризует соотношение занятых и свободных участков связывания при данной концентрации лиганда. Обычно для описания параметров связывания используют величину, обратную константе сродства,—
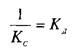
— константу диссоциации. Эта константа соответствует величине, при которой происходит насыщение 50% связывающих участков:
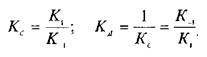
Если вместо константы сродства Кс использовать обратную ей величину Кд, то, подставив это значение в уравнение, характеризующее равновесную реакцию взаимодействия лиганда с рецептором, получим следующее уравнение:
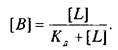
Приняв общее число рецепторов за 1, можно преобразовать уравнение к виду, аналогичному уравнению Михаэлиса, которое используется в энзимологии для описания кинетики обратимых ферментативных реакций:

где — концентрация комплекса фермент-субстрат; —
концентрация субстрата и Ks — константа диссоциации комплекса; — исходная концентрация субстрата.
Согласно этим уравнениям зависимость величины эффекта от дозы лиганда или фермента описывается гиперболой. Чаще всего для работы пользуются графическим выражением зависимости эффекта не от концентрации, а от логарифма концентрации лиганда. Графически зависимость результатов может быть представлена разными способами, однако наиболее информативным способом расчета являются координаты Скэтчарда. Действительно, помимо равновесной константы связывания и общей концентрации центров связывания этот метод позволяет определить концентрацию свободного лиганда, соответствующую данной концентрации комплекса В. Константа диссоциации равна котангенсу угла наклона прямой. Отрезок на оси абсцисс от точки пересечения с прямой до начала координат соответствует максимальному уровню насыщения центров связывания.

Таким образом, представление результатов равновесного связывания в координатах Скэтчарда дает информацию о характере протекающего процесса и позволяет определить важные параметры лиганд-рецепторного взаимодействия — константу диссоциации и концентрацию центров, способных образовывать комплексы с нейромедиатором.
В качестве примера изучения рецепторного связывания нейромедиатора с белковыми компонентами на мембране нейрона приведем экспериментальные исследования глутаматных рецепторов радиолигандным методом. Так, исследования параметров связывания Н-глутамата с синаптическими мембранами, выделенными из коры больших полушарий головного мозга крыс, показали их зависимость от чистоты материала, способов хранения, условий проведения реакции связывания и др. При стандартизации всех указанных условий зависимость специфического связывания Н-глутамата с синаптическими мембранами имеет насыщающий характер. Представление экспериментальных данных в координатах Скэтчарда свидетельствует о наличии на мембранах однородной популяции участков связывания с Кд - 89,4 нМ и Вмакс = 2,0 пмоль/мг белка.

Значение количества центров связывания, выраженное в СРМ, пересчитывается в фмоль/мг белка по следующей формуле: