Реферат: Мотонейрон, его строение и функции
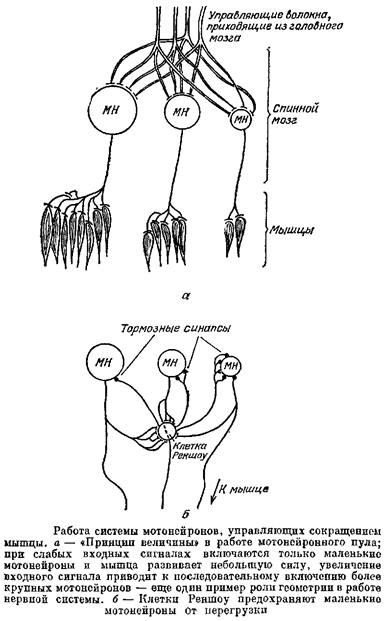
Чтобы этого не происходило, в МН-пуле есть специальное защитное устройство — так называемые клетки Реншоу. К этим клеткам идут специальные отростки аксоновМН, причем в основном от больших МН. Клетки Реншоу — тормозные нейроны, их аксоны оканчиваются на МН, причем преимущественно на маленьких. Когда сигнал, приходящий на пул сверху, велик, к маленьким МН приходит, с одной стороны, этот слишком большой для них сигнал, а с другой стороны — тормозной сигнал от клеток Реншоу; эти сигналы, имеющие разные знаки, складываются, и маленькие МН работают в нормальном для них режиме,
Частотное кодирование и нейроны без импульсов
У обычного нейрона имеется возбудимая мембрана, которая в ответ на входной сигнал, достигший порога1 выдает выходной сигнал в виде ПД всегда одной и той же формы и амплитуды. Таким образом, ПД не несет никаких признаков входного сигнала. Одна и та же клетка в нервной системе может генерировать импульс и в ответ на свет и в ответ на звук, и если мы будем регистрировать импульсы в ее аксоне, мы никаким способом не сможем узнать по какой причине они возникли
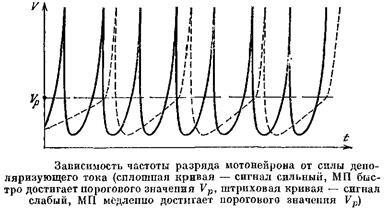
Получается, что нейрон при передаче сигналов «обедняет» информацию. Правда, оказывается, что хотя нейрон ничего не может сообщить об источниках входных сигналов, он может сообщить об их силе. Как это делается? Амплитуда входного сигнала — это сумма всех синаптических потенциалов, создаваемых активными синапсами. Пусть во время действия сигнала амплитуда его не меняется. Тогда через мембрану нейрона все время будет течь примерно постоянный синаптический ток. Легко сообразить, что чем больше ток, идущий через мембрану, тем с большей частотой будет работать нейрон. Действительно, после каждого очередного импульса мембрана нейрона гиперполяризуется, так как открываются калиевые каналы. Чем больше ток, текущий через мембрану нейрона тем быстрее он возвратит его потенциал к пороговому значению. Таким образом, чем больше амплитуда входного сигнала, тем выше частота выходных импульсов. Этот способ передачи информации называют частотным кодированием.
Однако, оказывается, существуют нейроны совершенно другого типа, резко отличающиеся по способу своей работы от «стандартных». Мембрана таких нейронов вообще невозбудима, т. е. не способна генерировать импульсы. Поэтому при поступлении входного сигнала на такой «безымпульсный» нейрон возникает не ПД определенной амплитуды, а только сдвиг мембранного потенциала, тем больший, чем больше сила сигнала.
Этот сдвиг потенциала электротонически передается по аксону к синапсам и вызывает там выделение медиатора. Если на клетку пришел большой внешний сигнал и вызвал сильный сдвиг потенциала, то медиатора выделяется больше и клетка сильнее воздействует на своего адресата. Таким образом, безымпульсные нейроны передают входной сигнал аналоговым способом: здесь все величины остаются непрерывными, не преобразуются в дискретные стандартные импульсы.
Где же в нервной системе используются такие безымпульсные нейроны? Прежде всего, таковы многие рецепторы, т. е. нервные клетки, которые реагируют на сигналы из внешней среды. Например, безымпульсные нейроны глаза усоногого рака меняют под действием света свой МП непрерывным образом и передают эти изменения электротонически на расстояние в несколько сантиметров. Рецепторы глаз человека и других позвоночных — палочки и колбочки — тоже являются безымпульсными нейронами, Другие безымпульсные нейроны сами не являются рецепторами, но собирают сигналы от рецепторов и передают их дальше аналоговым образом. Такой нейрон обнаружен, например, у речного рака; он собирает сигналы от нескольких механорецепторов, и сдвиг его потенциала пропорционален скорости движения воды относительно тела, когда вода течет от хвоста к голове. Почему именно в таком направлении? Потому, что рак пятится хвостом вперед, и эти безымпульсные нейроны помогают ему измерять скорость его попятного движения. В сетчатке человека и животных безымпульсными являются не только палочки и колбочки, но и многие другие типы клеток, которые получают сигнал от фоторецепторов. Вся обработка сигналов в сетчатке в сложной сети этих разных типов клеток осуществляется без импульсов, аналоговым образом. Генерируют импульсы только выходные клетки сетчатки — ганглиозные клетки, которым нужно передать сигнал от сетчатки в мозг по длинным аксонам, образующим зрительный нерв,
Клетки-генераторы и мозаика каналов
Мы видели, что для генерации ритмических движений при плавании в нервной системе пиявки имеется специальная схема из нейронов, которая генерирует периодические сигналы. Оказывается, источником периодических сигналов может быть и одиночный нейрон. Это еще один очень важный тип «нестандартных» нейронов, часто тоже безымпульсных. Эти нейроны способны сами по себе работать ритмически: потенциал такого нейрона все время колеблется, при деполяризации концевые веточки его аксона выделяют больше медиатора, и он возбуждает обычные импульсные нейроны, а при гиперполяризации выделение медиатора прекращается.
Эти нейроны играют важную роль в управлении ритмическими движениями: ходьбой, полетом, дыханием и т. д. Первый такой нейрон, управляющий вентиляцией жабер краба, был открыт в 1971 г. А в 1975 г. был открыт второй нейрон такого типа, управляющий передвижением таракана. Потом были описаны нейроны, с помощью которых поют цикады или работает желудочная мельница омара. Недавно шведский ученый Стен Гриллнер обнаружил периодически работающие безымпульсные нейроны, которые обеспечивают плавание миноги. Это значит, что и у позвоночных спонтанно активные безымпульсные нейроны могут управлять движениями.
Вопрос о том, как возникает ритм в нейронах-генераторах, не имеет однозначного ответа. Прежде всего, существует несколько принципиально разных способов организации такой ритмики. И первый из них — поместить в мембрану такого нейрона подходящий набор ионных каналов. Однако даже и в этом варианте задача не становится однозначной: экспериментально показано, что в разных клетках ритмические колебания потенциала создаются разными наборами ионных каналов.
Рассмотрим один из примеров ионного механизма генерации таких потенциалов. Пусть у нас есть клетка, ПП которой равен —60 мВ; нас не интересует, за счет каких каналов он создается, мы будем считать, что их относительно мало и они создают относительно небольшие токи.
Пусть, кроме того, в мембране этих нейронов есть еще два типа каналов: кальциевые и калиевые. Кальциевые каналы открываются при деполяризации и сами по себе не закрываются, не ипактивируются; чтобы они закрылись, надо либо гиперполяризовать клетку, либо накопить в ней достаточно много кальция. Калиевые же каналы устроены так, что они открываются под влиянием кальция, действующего с внутренней стороны мембраны, если его концентрация достигает некоторого порогового уровня. Пусть, наконец, ПП в —60 мВ является пороговым для кальциевых каналов.
Такой нейрон будет работать следующим образом. При МП в —60 мВ кальциевые каналы откроются и ионы кальция начнут поступать в клетку? которая деполяризуется, и это состояние будет некоторое время сохраняться. Когда внутри клетки накопится достаточное количество Са, кальций-чувствительные К-каналы откроются, и возникнет идущий наружу калиевый ток. Этот ток гиперполяризует нейрон и Са-каналы закрываются.
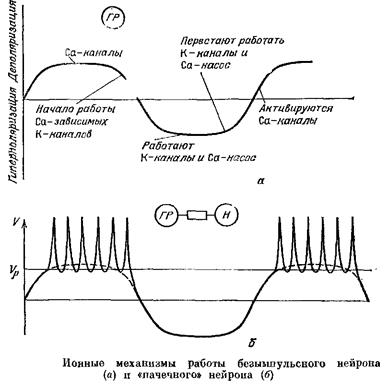
Са-насосы постепенно снижают концентрацию Са внутри клетки, интенсивность их работы снижается, и МП будет постепенно приближаться к «ПП»„ При достижении этого уровня вновь откроются Са-каналы и цикл повторится.
Длительность цикла такого нейрона может быть разной в зависимости от чувствительности разных типов каналов и от плотности зтих каналов, обычно цикл довольно велик. Нейроны работают с тем периодом, который характерен для управляемого ими движения. Например, бесспайковый нейрон таракана работает в ритме ходьбы и управляет сгибанием и разгибанием ног.
Описанный нами нейрон можно «остановить». Если на него будут непрерывно действовать тормозные синапсы, то нейрон будет гиперполяризован, а его Са-каналы все время закрыты. Возможен и «обратный» вариант: если ПП такого нейрона будет равен не —60 мВ, а, например, — 70 мВ8 то нейрон будет «молчащим», пока на него не подействуют возбуждающие синапсы, которые сдвинут его МП до —60 мВ. Тогда в нейроне и начнутся ритмические колебания потенциала. Для управления ходьбой или полетом используются именно такие нейроны, в которых ритмические колебания потенциала могут запускаться и останавливаться. Ведь не может же насекомое всю жизнь безостановочно бегать или летать!
Нейроны-гибриды
Давайте решим такую задачу. Пусть есть два нейрона: первый—генератор ритма, а второй — стандартный, с обычной возбудимой мембраной. Соединим их электрическим синапсом. Как будет работать эта схема?
Легко понять, что как только МП первого нейрона достигнет порога возбуждения, второй нейрон выдаст импульс, обычный ПД, стандартной амплитуды и длительности. Но период работы клетки-генератора гораздо больше, чем длительность ПД. Кончится и рефрактерный период, а генератор будет продолжать передавать на второй нейрон потенциал, достаточный для его возбуждения,— возникнет новый импульс и т.д. Получается, что во время деполяризации клетки-генератора на мембране второй клетки успевает возникнуть целая серия ПД, а во время гиперполяризации получается пауза между сериями.
Оказывается, такая схема существует в действительности и в очень интересном воплощении: в одной и той же клетке. В мембране такой клетки есть и каналы, обеспечивающие возникновение ПД, и весь механизм для генерирования медленного ритма. Такой «гибрид» стандартного нейрона и пейсмекера называют «пачечным» нейроном, потому что он периодически выдает «пачки», т. е. серии импульсов, разделенные паузами.
Еще одна задача. В мембране пачечного нейрона соединены два набора каналов: от генератора ритма и от обычного нейрона. А теперь представим себе клетку-генератор, из мембраны которой удалены калиевые каналы, открывающиеся под действием кальция.
Легко понять, что такая клетка уже не будет генерировать колебания: если мембрану деполяризовать настолько, чтобы открылись Са-каналы, то они все время остаются открытыми на некотором новом равновесном уровне. Получается, что у клетки будут два устойчивых уровня МП: один — тот, к которому она приходит при закрытых Са-каналах, а другой — тот, который устанавливается при работе этих каналов. Такой нейрон можно перевести из одного состояния в другие, действуя на него либо возбуждающими, либо тормозными синапсами.
Оказывается, что и такие «бистабильные» клетки тоже встречаются в действительности.
Итак, в зависимости от того, какой набор ионных каналов есть в мембране, можно получить обычный нейрон, генерирующий ПД, можно получить безымпульсный нейрон, можно получить «пачечный» нейрон, «бистабильный» нейрон и другие варианты. Подобно тому, как ребенок выкладывает из мозаики разные картинки, природа с помощью разного набора ионных каналов обеспечивает разнообразие свойств нейронов, что и позволяет им выполнять разные функции.
Мы видели, что сами каналы состоят из частей: «трубка», «ворота», «реле». Видимо, как это предполагает П.Г. Костюк, они могут соединяться в разных сочетаниях, давая каналы с разными свойствами. Точно так же на следующем уровне каналы, собираясь в разных сочетаниях в мембране одного нейрона, могут определять его свойства. Наконец, из нейронов с разными свойствами образуются разные нейронные сети. Природа широко использует метод «блочного строительства».
А теперь вернемся еще раз к нейронам-генераторам. Мозаика ионных каналов — это один из способов создать колебания потенциала. Возможен и другой. Сейчас открыты колебательные химические реакции, во время которых компоненты реакции периодически меняют свою концентрацию. В открытии этих реакций и в том, что они получили широкое признание, важную роль играли советские ученые Б.П. Белоусов, А.М. Жаботинский и С.Э. Шноль. Представьте себе, что один из компонентов такой колебательной реакции умеет открывать ионные каналы и деполяризовать клетку. Это будет принципиально другой способ работы нейронов-генераторов, так как тут автоколебательная система находится не в мембране клетки, а в цитоплазме, и образована не совокупностью ионных каналов, а цепью химических реакций. Этот способ генерации колебаний интересен тем, что он связывает электрические мембранные процессы с биохимией клетки, с обменом веществ.
А теперь о сердце
Если вы спросите у биолога: «Что заставляет сердце биться?», то он ответит вам вопросом на вопрос: «У какого животного?», и неспроста. Оказывается, у разных животных сердечный ритм создается по-разному. Наиболее естественно предположить, что «водителями» сердечного ритма являются уже знакомые нам клетки-генераторы. И действительно, например, у краба или омара есть специальный нервный ганглий из 9 клеток, управляющий сокращениями сердца. Среди этих клеток есть клетки-генераторы, почти такие же, как те, о которых мы только что рассказывали. Особенность клеток сердечных ганглиев состоит в том, что в их аксонах есть каналы, т. е. мембрана их аксонов возбудима. Это похоже на выдуманную нами «гибридную схему», причем аксон играет роль клетки 2. Когда тело клетки сердечного ганглия деполяризовано, в аксоне возникает пачка импульсов, вызывающая сокращение сердечной мышцы. Без этих нервных клеток сердце краба останавливается.