Реферат: Мутационная изменчивость микроорганизмов – производителей антибиотиков
У грибов существуют разнообразные типы скрещивания, которые используются в генетических исследованиях. Многие грибы-аскомицеты и базидиомицеты обладают сложноорганизованными системами скрещивания, препятствующими самооплодотворению и другими формами инбридинга. Половой процесс контролируется системой несовместимости. У некоторых грибов система несовместимости биполярна; при этом процесс скрещивания контролируется всего одним локусом, который существует в двух альтернативных аллельных формах. У других грибов (например, Schizophyllum commune) система несовместимости тетраполярна. У них тип спаривания определяется двумя генами, каждый из которых имеет множество аллельных форм. Для успешного скрещивания два партнера должны обладать разными аллелями каждого из двух локусов типа спаривания.
2. Мутагенез и методы выделения мутантов
Генетическое изучение микроорганизмов, создавшее фундамент для современной селекции, стало возможным только, когда были разработаны способы выделения клоновых культур, или клонов.
Клон — это генетически однородное потомство одной клетки, например колония, возникшая из одной клетки при рассеве культуры на плотной питательной среде. Исследуя свойства такой колонии, можно получить представление и о признаках породившей ее клетки.
Важно отметить, что при последующих пересевах клоновой культуры в результате процесса изменчивости могут появиться варианты, отличающиеся от исходного. И тогда клоновая культура клеток превращается в генетически разнородную клеточную популяцию. Клоновая по происхождению культура, наследственная однородность которой поддерживается отбором по специфическим признакам, называется штаммом. Получение и поддержание высокопродуктивных штаммов — основная задача селекционной работы.
Важнейшим методом селекции микроорганизмов является отбор мутантов, т. е. организмов с измененными наследственными признаками, которые появляются в результате мутаций. В самом широком смысле мутацию можно определить как внезапно возникающее наследуемое изменение в генетическом материале клетки. Следует различать мутации цитоплазматические, затрагивающие внехромосомные генетические детерминанты, и ядерные, или хромосомные.
В свою очередь, хромосомные мутации можно разделить на три основных типа: 1) изменение числа хромосом; 2) изменение числа и порядка расположения генов (перестройки хромосом или структурные изменения); 3) изменения индивидуальных генов (внутригенные изменения, или мутации в наиболее узком смысле этого слова) (Ш. Ауэрбах, 1978). В селекции микроорганизмов основное значение имеют последние два типа мутаций.
Хромосомные перестройки включают: выпадения участков хромосомы (делении), удвоения (дупликации) или умножения (амплификации) числа отдельных генов или группы генов, вставки участков хромосом в новые места (транспозиции), обмен участками между хромосомами (транслокации), изменения порядка расположения генов на хромосоме (инверсии). Такие мутации могут вызывать как утрату функций, так и приобретение новых признаков, в частности в связи со слиянием генов, которые могут оказаться под контролем несвойственных им регуляторных элементов. При этом могут появиться гибридные белки, увеличиться (уменьшиться) количество продуктов определенных генов. За исключением амплификации, псе хромосомные перестройки стабильны.
Внутригенные мутации изменяют последовательность оснований ДНК в пределах одного гена. Это могут быть выпадения или вставки одного или нескольких оснований, нарушающие порядок считывания гена в процессе трансляции. В клетках такого типа мутанта синтезируется неактивный белок с измененной последовательностью аминокислот. При транзициях происходит замена какого-либо одного пурина или пиримидина (тимина или цитозина) на другой пурин или пиримидин соответственно. При трансверсиях пуриновые основания заменяются на одно из двух пиримидиновых и, наоборот, пиримидиновое основание — на одно из двух пуриновых.
Важной характеристикой мутантов является их способность, к реверсии, т. е. обратному мутированию к исходному фенотипу. Мутанты, которые появляются в результате реверсии, называются ревертантами. При истинных обратных мутациях в ДНК восстанавливается исходная последовательность оснований. Так ревертируют точковые мутации — замены оснований, вставки или выпадения одного или нескольких нуклеотидов.
Кроме того, реверсии могут произойти благодаря супрессорным мутациям. При внутригенной супрессии вторая мутация возникает в том же гене, что и первичная мутация, и приводит к более или менее полному восстановлению функции белка. При внегенной супрессии вторая мутация затрагивает другой ген. Так, ошибки кодирования, связанные с нонсенс-мутациями и некоторыми мутациями со сдвигом рамки, могут частично исправляться мутациями в генах, кодирующих РНК Восстановленная активность поврежденного белка при этом обычно не превышает 10 % от исходного уровня.
Когда невозможно провести прямой отбор мутантов, исследуют колонии на индикаторных чашках, применяют тесткультуры микроорганизмов или перепечатывают колонии на различные среды, т. е. используют метод отпечатков, или реплик. Иногда приходится выращивать каждую колонию и определять в культуральной жидкости интересующую активность. Индикаторные чашки дают возможность различать мутанты по цвету колоний и проводить тестирование разнообразных фенотипов в больших популяциях. Такие чашки могут содержать среды с индикатором, выявляющим различие в рН между теми колониями, которые метаболизируют определенные углеводы, и теми, которые не обладают такой способностью. Так, на агаре с трифенилтетразолием колонии, не сбраживающие лактозу, приобретают ярко-красный цвет, а колонии, сбраживающие этот дисахарид, остаются неокрашенными. Используются также специально приготовленные субстраты, распадающиеся с образованием красителя при их гидролизе ферментами, наличие которых тестируется на этих чашках. Иногда вносят субстраты, изменяющие прозрачность сред, и наблюдают образование вокруг колоний мутантов зон просветления.
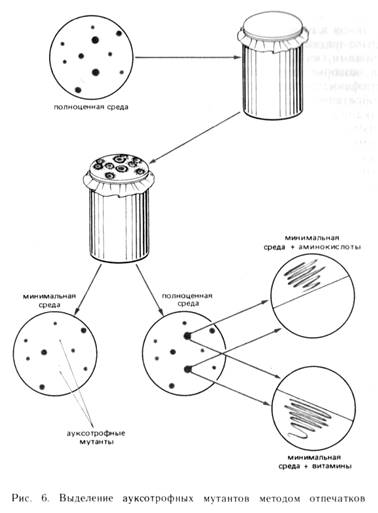
В 1952 г. Д. Ледерберг и Э. Ледерберг ввели для непрямого отбора мутантов метод отпечатков (рис. 6). Согласно этому методу, чашки Петри засеваются с таким расчетом, чтобы на каждой из них выросло 50—200 колоний. Стерильный бархат или фильтровальную бумагу натягивают на металлический или деревянный цилиндр и закрепляют металлическим кольцом. Чашки с выросшими колониями переворачивают и прикладывают к бархату. Затем к этому же бархату (с отпечатками колоний на нем) прикладывают чистые чашки с различными средами. После соответствующей инкубации на них образуются колонии в том же расположении, что и на исходной (матричной) чашке. Если матричная чашка содержала полноценную среду, то отпечатками на минимальные среды можно выявить ауксотрофные мутанты. Различные модификации этого метода широко используются для выделения мутантов с питательными потребностями, а также мутантов, чувствительных к различным физическим, химическим и биологическим агентам (температура, антибиотики, фаги и др.).
Получение ауксотрофных мутантов с помощью метода отпечатков может быть облегчено, если обогащать ими популяцию клеток, используя пенициллин или его аналоги. Эти антибиотики убивают только растущие клетки. Например, ауксотрофные по треонину мутанты Е. coli не растут на минимальной среде без треонина. В этих условиях пенициллин будет убивать только прототрофные клетки. Однако полной селекции ауксотрофов эта процедура не обеспечивает.
Иногда приходится проводить несколько циклов обработки пенициллином, попеременно выращивая бактерии в среде с треонином без пенициллина и в минимальной среде без треонина с пенициллином. После одного цикла достигается обогащение желаемыми мутантами в 1000 и даже в 10000 раз. Следовательно, если необходимая мутация возникает спонтанно с частотой 10~7 , то достаточно просмотреть несколько тысяч колоний, выживших после обработки пенициллином, чтобы найти колонию с нужной мутацией.
Вывод
Сегодня в этой области произошли явные перемены: если раньше единственным используемым генетическим методом был отбор улучшенных штаммов, то сегодня предлагаются совсем новые подходы, основанные на технологии рекомбинантных ДНК (генетическая инженерия). С их помощью путем ферментации можно получать новые виды продукции: белки и пептиды человека, антигены вирусов. Надо сказать, что большой интерес к биотехнологии в значительной мере обусловлен именно появлением генетической инженерии. Сейчас широкое применение находят методы модификации генетического материала как in vivo, так и in vitro для разработки новых и модернизации существующих биотехнологических процессов.
Литература
1. Биотехнология Принципы и применение Пер с англ. /Под ред. И. Хтгенса, Д.Беста, и Дж.Джонса. М.: Мир, 1988.-480ст.
2. Экологическая биотехнология. Под. ред. Форебера и Дис.Вайза. – Л.: Химия. – 1991 – 120с.
3. Герасименко В.Г. Биотехнология. – Киев.: Высшая школа – 1989. - 342 с.
4. Белков В.А. Биотехнология. Микробиологическое производство биологически активных веществ. – М.: Высшая школа. – 1997. – 220 с.
5. Мильчук М.Д., Новак Т.В., Кунах В.А. Біотехнологія рослин. – Київ, Поліграфконсалтінг, 2003. – 250 с.
6. Рыльский А.Ф. Краткий курс лекций по биотехнологии (для студ. Дневного отделения биологического факультета).- Запорожье: ЗГУ, 1996.- 75с.