Реферат: От чего зависит судьба гена
В.Л. Карпов, доктор биологических наук, заместитель директора Института молекулярной биологии им.В.А.Энгельгардта РАН.
Все клетки любого многоклеточного организма содержат одинаковую генетическую информацию, но во время его развития она считывается избирательно, одновременно все гены никогда не работают. Например, в клетках печени активны только те гены и синтезируются только те белки, которые нужны для ее функционирования, в клетках почек работают лишь гены, необходимые для выполнения функций этого органа, и т.д.
За счет чего же стабильно включаются и выключаются определенные гены и не теряется при этом остальная генетическая информация?
Оказалось, что избирательность считывания контролируют особые - эпигенетические - механизмы (греч. epi означает над, сверх). Наследование этого контроля называют эпигенетическим. Оно не затрагивает информацию, записанную в ДНК, тут действуют другие, “надгенетические”, механизмы. Именно они определяют будущее клетки - какой ей быть, какие гены включать для синтеза белков, а какие - выключать.
Первая ступень контроля
Существует несколько уровней контроля активации генов. Один из них - модификация ДНК. Подобному изменению подвергаются гены, которые необходимо выключить в конкретном типе клеток, и оно должно быть таким, чтобы ДНК не теряла способности копироваться при делении клетки. Тогда эта модификация сохранится и в дочерней клетке, и в ней ген тоже будет выключен. В настоящее время известна только одна модификация ДНК, не нарушающая кодирование и копирование. Это - метилирование, присоединение к одному из четырех азотистых оснований, цитозину, метильной группы (–СH3). Метилируется при этом не любой цитозин, а лишь тот, что находится в составе динуклеотида CG. В ДНК позвоночных животных такие динуклеотиды встречаются нечасто, и более половины их бывают метилированы специально предназначенным для этого ферментом - метилазой. Важно, что СH3-модификация копируется при удвоении ДНК. Если на старой цепи ДНК есть метилированные цитозины, фермент присоединяет метильные группы к таким же основаниям и на вновь синтезируемой цепи. Благодаря этому модификация передается в клеточных поколениях, т.е. сохраняется информация о том, что данный ген должен быть выключен.
Каким же образом действует модификация на ген? По-видимому, существует два основных механизма с противоположным действием: отталкивание белков, активирующих ген, и наоборот, привлечение к метилированному фрагменту белков, участвующих в очень плотной упаковке модифицированного гена за счет изменения структуры хроматина. Хроматин представляет собой чрезвычайно сложный и динамичный комплекс, состоящий главным образом из ДНК и белков. На уровне хроматина и осуществляется второй эпигенетический механизм контроля активности генов.
Не только для упаковки ДНК
Молекула ядерной ДНК, как известно, отнюдь не мала, ее линейный размер, например в клетке человека, достигает почти двухметровой длины, тогда как диаметр ядра не превышает 0.01 мм. Столь длинная ДНК упакована в нем таким образом, что из нее в нужный момент и в нужной комбинации извлекается необходимая информация. Совмещение плотной упаковки ДНК и избирательного извлечения из нее генетической информации обеспечено хроматином. Можно сказать, что именно он служит носителем генетической информации в клетках высших организмов, так как от него зависит избирательность работы генов. Огромное разнообразие фенотипического проявления наследственного материала - результат подобной избирательности и комбинаторики. Не удивительно поэтому, что почти при 80-процентной схожести геномов человека и мыши сами организмы сильно отличаются друг от друга. В настоящее время уже установлено, что на уровне хроматина, а не ДНК, наиболее эффективно работают многие ферменты, участвующие в избирательном считывании генетической информации, ее удвоении при делении клетки, исправлении возникающих ошибок и т.д.
Термин “хроматин” введен в научную лексику в 1888 г. Им стали называть окрашиваемое содержимое ядра, обнаруженное за восемь лет до этого. Истинная роль хроматина в жизнедеятельности клетки начала проясняться почти 100 лет спустя, после открытия в 1973-1974 гг. нуклеосомы - главного структурного элемента хроматина. Ученые пришли к выводу, что она представляет собой ту единицу, на основе которой строится сложнейший механизм, предназначенный для дифференциального включения генов в разных клетках и на разных стадиях индивидуального развития организма. Чтобы понять, почему одни гены работают в каком-то типе клеток, а другие - нет, и как целенаправленно управлять этим процессом, нужно обратиться к структурной организации хроматина.
Гистоны и нуклеосома
Здесь уже упоминалось, что хроматин - это очень сложный комплекс ДНК и белков. Большинство их составляют гистоны, обнаруженные в 1884 г. немецким биохимиком А.Косселем через 15 лет после открытия нуклеина (ДНК) И.Ф.Мишером, швейцарским биохимиком и физиологом. Гистоны оказались универсальными компонентами хроматина. По массе они приблизительно равнялись ДНК, и некоторое время их считали носителями генетической информации. Но потом отвели роль регуляторов генной активности, однако позже выяснилась необоснованность этой точки зрения: огромное разнообразие гистонов, на которое она опиралась, было вызвано методом их выделения - эти белки интенсивно разрушались. В конце концов метод экстракции удалось усовершенствовать, и разнообразие гистонов свелось к пяти основным типам: Н1, Н2А, Н2В, Н3 и Н4 (Н - от англ. Нistone).
Гистоны - белки небольшой молекулярной массы - относятся к наиболее консервативным по первичной структуре белкам в ряду от простейших эукариот до высших. Например, аминокислотная последовательность Н4 из вилочковой железы теленка и проростков гороха отличается только двумя остатками из 102, составляющих всю молекулу. Характерное свойство гистонов - высокое содержание лизина и аргинина, положительно заряженных аминокислотных остатков. Они находятся преимущественно в аминной (N-) и карбоксильной (С-) концевых областях белка, называемых “хвостами”. Их функция долгое время оставалась непонятной, и существовало даже мнение, что они не очень важны для нормальной жизни клетки. Однако, как выяснилось позже, именно гистоновые “хвосты” играют главную роль в эпигенетических механизмах. В центральных, самых консервативных, участках полипептидной цепи гистонов преобладают остатки гидрофобных аминокислот. Именно эти центральные области участвуют в образовании специфических комплексов из молекул гистонов: тетрамера (Н3)2-(Н4)2 и двух димеров Н2А-Н2В. Из них формируется нуклеосомная сердцевина, ядро (англ. core), на которое и навивается ДНК.
В основе формирования нуклеосомы лежат несколько принципов, сформулированных А.Корнбергом [1]. В число восьми гистоновых молекул, составляющих нуклеосомное ядро, входят по два Н2А и Н2В с высоким содержанием лизина и по два Н3 и Н4, богатых аргинином. На этот октамер, как на катушку нитка, наматывается ДНК, образуя левозакрученную суперспираль с шагом 28 A. Навиться может 1.75 витка (по 80 пар нуклеотидов в одном) или 2-2.5 витка. Первую нуклеосому называют минимальной, вторую - полной. Они отличаются количеством не только ДНК, но и гистонов: в минимальной нуклеосоме, как уже сказано, содержится восемь молекул гистонов, а в полной к ним добавляется еще один - Н1, который связывается с межнуклеосомной ДНК.
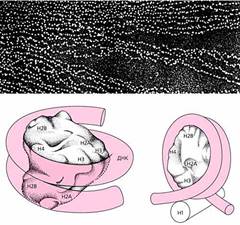
Электронная микрофотография хроматина (вверху) и модели минимальной (внизу слева) и полной нуклеосомы.
На фотографии хроматин виден как нить с нанизанными на нее бусинками. Ядро нуклеосомы, или минимальная нуклеосома, состоит из восьми молекул гистонов - по две молекулы каждого вида: Н2А, Н2В, Н3 и Н4. Этот октамер обвит сегментом ДНК, делающим вокруг ядра 1.75 оборота (внизу слева), а гистон-1 (Н1) в полной нуклеосоме “сшивает” ДНК в начале и в конце обвитого сегмента (внизу справа).
В нуклеосоме гистоны взаимодействуют с ДНК строго определенным образом. В начале 80-х годов в лаборатории А.Д.Мирзабекова был разработан метод анализа ДНК-белковых взаимодействий, основанный на сшивках этих молекул, с помощью которого удалось расшифровать порядок расположения гистонов в нуклеосоме [2]. Оказалось, что он одинаков в организмах, принадлежащих не только к разным видам, родам и классам, но даже царствам живой природы - грибам, растениям и животным. Это открытие объяснило высокую консервативность гистонов, необходимую для формирования столь сложного комплекса, какой представляет собой нуклеосома.
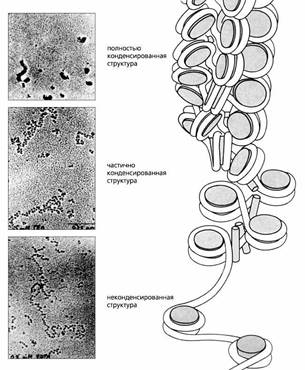
Хроматин в разной степени конденсации - на микрофотографии и на схеме.
В растянутой форме (нижняя часть рисунка) хроматин имеет вид нити с бусинками-нуклеосомами. За счет гистона-1 (Н1) он частично уплотняется, конденсируется (в середине), а затем конденсируется полностью (механизм остается неизвестным), и гены в этой части ДНК совершенно неактивны.
Большой вклад в формирование идей о нескольких уровнях упаковки ДНК в составе хроматина внесли работы Г.П.Георгиева и Ю.С.Ченцова [3]. Первый уровень - образование фибриллы диаметром 100 A; в ней ДНК становится компактнее в 6-7 раз. Далее при участии гистона-1 формируется 30-нанометровая фибрилла, в результате ДНК уплотняется еще в 20-50 раз. Существует огромное количество версий, объясняющих механизмы этих процессов, но до сих пор нет единого мнения на этот счет. Еще меньше известно об упаковке ДНК в петли и домены. Однако ни у кого нет сомнений в том, что структура нуклеосомы играет главную роль в укладке ДНК на всех уровнях. Именно взаимодействия между нуклеосомами определяют степень компактности ДНК.

Структура минимальной нуклеосомы (вид вдоль оси суперспирали ДНК). Разрешение 2.8 A
Показана половина нуклеосомы с одним витком суперспирали (73 пары оснований) и четырьмя молекулами гистонов (Н2А, Н2В, Н3, Н4). Их спиральные сегменты изображены в виде цилиндров, неструктурированные участки между ними - в виде петель, буквами N и С обозначены “хвосты”, т.е. аминные и карбоксильные концы молекул. Места, в которых ДНК контактирует с гистонами, указаны белыми крюками.
Рентгеноструктурный анализ, позволяющий получать данные о пространственном расположении атомов в кристалле, для нуклеосомы особенно сложен в силу того, что этот комплекс состоит из многих компонентов. Но в конце 90-х годов в лаборатории Т.Ричмонда такой анализ был успешно осуществлен и получено “изображение” структуры нуклеосомы с высоким разрешением - около 2.8 A [4]. Таким образом, появилась основа для более глубокого понимания тех механизмов, которые обеспечивают участие нуклеосомы в уплотнении ДНК и процессах регуляции генной активности на уровне хроматина.
Хвосты, хвосты, хвосты
Нуклеосома формируется по принципу многоуровневого узнавания, обусловленного строением гистонов. Молекула любого из них содержит центральный структурированный трехспиральный домен и неструктурированные N- и C-“хвосты”. Гистоны попарно узнают друг друга, и в этом важна роль особой гистоновой укладки. Спиральные домены взаимодействуют между собой, образуя структуры, названные рукопожатием, в результате чего возникают гетеродимеры - один Н3-Н4 и два Н2А-Н2В. Из первого димера в свою очередь образуется тетрамер (Н3)2-(Н4)2. Таким образом, этот тетрамер и два димера Н2А-Н2В составляют гистоновый октамер, сердцевину нуклеосомы.
Интересно, что мотив гистоновой укладки обнаружен во многих белках, которые активируют или подавляют считывание информации с ДНК. Из этого следует, что между структурой хроматина и механизмом транскрипции существует эволюционная связь.
Формой гистоновый октамер напоминает клин, благодаря чему обеспечивается левое закручивание спирали ДНК. На поверхности октамера, обращенной к ДНК, из центральных доменов формируются особые структурные элементы, или мотивы, которые можно разделить на три основных типа. Мотивы первого типа образованы -мостами - спаренными петлями гистоновых димеров. Мотивы второго типа - это спаренные N-концевые сегменты длинных спиральных доменов каждого гистона, образующего гетеродимер. Наконец, третий тип составляют два мотива из двух дополнительных a-спиральных участков гистона-3, расположенных вдоль суперспирали ДНК в месте ее входа в нуклеосому и выхода из нее. Поскольку перечисленные структурные элементы содержат положительно заряженные аминокислоты (в основном аргинины), октамер электростатически взаимодействует с отрицательно заряженными фосфатными группами сахарофосфатного остова ДНК. Таким образом, 14 малых бороздок ДНК втягивают в себя 14 содержащих аргинин мотивов гистонового октамера, расположенных на его поверхности. В результате ДНК жестко закрепляется почти независимо от нуклеотидной последовательности. Благодаря этому обеспечивается универсальность уплотнения ДНК на октамерах.
Итак, за счет консервативной центральной части гистонов формируется сам октамер и закрепляется ДНК, чем обеспечивается первый уровень ее уплотнения. ДНК оказывается на поверхности октамера и остается доступной для взаимодействия с другими белками.
Зачем же нужны гистоновые “хвосты”? Чтобы понять это, обратимся снова к нуклеосоме. Судя по рентгеноструктурному анализу, все они выходят на ее поверхность, причем “хвост” гистона-3 особенно далеко простирается за пределы нуклеосомы.
“Хвосты” гистонов очень подвижны. Выступая поверх не только нуклеосомы, но даже хроматиновой фибриллы, они участвуют в межнуклеосомном взаимодействии и подвергаются многочисленным модификациям. Среди них можно отметить ацетилирование, фосфорилирование, метилирование, AДФ-рибозилирование и присоединение белка убиквитина [5]. Эти модификации изменяют заряд, гидрофобность и другие свойства поверхности белковых глобул. Гистоновые “хвосты”, расположенные снаружи хроматиновой фибриллы, составляют до 25-30% от массы индивидуальных гистонов и различными заряженными группами мозаично “раскрашивают” поверхность монотонного хроматина. Кроме того, специализированные модифицирующие ферменты могут изменять эту мозаику, сочетая разные модифицирующие группы. Так формируется сложная матрица, которую узнают другие регуляторные белки, внешние сигналы. Более того, поскольку концевые домены гистонов участвуют и в межнуклеосомном взаимодействии, за счет модификаций хроматиновая фибрилла разрыхляется или, наоборот, уплотняется. Это, в свою очередь, облегчает или затрудняет доступ многочисленных регуляторов к ДНК.
--> ЧИТАТЬ ПОЛНОСТЬЮ <--