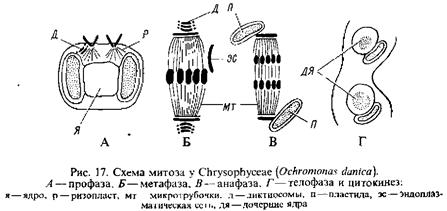
Реферат: Отдел золотистые водоросли
Род хромулина, как и охромопас, представлен одиночными голыми клетками, но с одним жгутиком.
Род уроглена (Uroglena). Виды этого рода обычны в пресноводном планктоне озер и прудов, представлены шаровидными или эллипсоидальными слизистыми колониями, в которых протопласты находятся на концах слизистых.
Семейство динобриевые объединяет свободно живущие или прикрепленные, одиночные или колониальные формы, клетки которых заключены в домик – мертвое, выделяемое протопластом образование. Типичный представитель – род динобрион, виды которого широко распространены в пресноводном планктоне и при массовом развитии нередко вызывают цветение воды в озерах и прудах. Голая, одетая цитоплазматичес кой мембраной клетка заключена в бокаловидном домике, образованном целлюлозными микрофибриллами с петлеобразным расположением; базальньгй конус домика состоит из продольно ориентированных микрофибрилл. В состав домика входит и большое количество аминокислот. Протопласт прикреплен к одной стороне основания домика базальной ножкой, способной к удлинению, сокращению или закручиванию, что позволяет клетке двигаться внутри домика: в этой части клетки находятся многочисленные микротрубочки. Клетка овальных очертаний содержит одно ядро, расположенное между хлоропластами. Два хроматофора различаются по длине, меньший располагается сзади. В хлоропласте можно видеть тилакоиды, жировые капли. На переднем конце более крупного хроматофора имеется глазок, состоящий из рядов электронно-плотных гранул несколько различной величины. В цитоплазме находятся эндоплазматический ретикулум, полирибосомы, митохондрии, в передней трети клетки - одна крупная диктиосома из приблизительно 12 диктиосомальных цистерн, от которых по краям отшнуровываются многочисленные пузырьки. В различных местах цитоплазмы содержатся свободные липидные гранулы, в задней части клетки – крупная вакуоль с хризоламинарином. Клетка имеет два жгутика: длинный и более короткий. Проксимальная часть короткого гладкого жгута несет вздутие, расположенное в непосредственной близости от стигмы. От базального тела к ядерной оболочке тянется поперечно исчерченный корень, кроме того, имеются 7 микротрубчатых корней, распространяющихся на периферию клетки. Клеточное тело соприкасается со стенкой домика не только базальной ножкой, но и сравнительно широкой экваториальной областью, в которой в молодых и делящихся клетках наблюдаются обильные пузырьки – дериваты аппарата Гольджи. Это заставляет предположить, что материал домка секретируется именно этими пузырьками. При продольном делении клетки один из дочерних протопластов или оба покидают материнский домик и, оседая у его края, вращаясь вокруг своей оси, формируют новый домик. Таким образом, возникают древовидные колонии различной формы. При образовании статоспоры базальная ножка, по-видимому, отделяется от стенки домика, и клетка мигрирует к его отверстию, где округляется, теряет жгутики и образует цисту или статоспору. По крайней мере часть статоспор Dinobryoncylindricum – результат гологамного полового процесса.
Семейство Синуровые – Synuraceae включает одноклеточные и колониальные формы, у которых клетки кнаружи от цитоплазматической мембраны несут покров из окремнелых чешуек.
Род малломонас (Mallomonas), обычный в пресноводном планктоне, наиболее богат видами. Клетки различной формы покрыты окремнелыми чешуйками или чешуйками со щетинками. В качестве представителя рода можно рассмотреть один из наиболее крупных видов – Mallomonaseaudata, для которого подробно описана не только ультраструктура содержимого клетки, чешуек и щетинок, но и механизм их образования, освобождения и отложения на поверхность клетки. Такого рода исследований сравнительно немного.
М. caudataимеет два жгутика, из которых в оптическом микроскопе различим только один'. Этот жгутик нормального строения и несет два ряда волосовидных мастигонем. Второй жгутик, не различимый в световом микроскопе, выступает из клетки на короткое расстояние и скрыт покровом чешуек; он обладает аксонемой 9 – 9 + 0.
 | 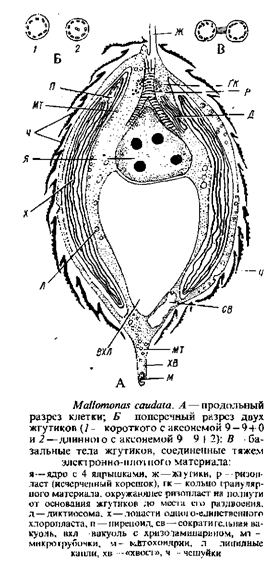 |
Два базальных тела соединены тяжем электронно-плотного материала. Ризопласт соединяет базальные тела с ядром, делясь непосредственно над ядром на две ветви. Несколько выше разветвления поперечно исчерченный корень окружен кольцом гранулярного материала, имеются и микротрубчатые корни. Одно ядро, содержащее до четырех ядрышек и расположенное по соседству с основаниями жгутиков, окружено оболочкой из двух мембран; наружная мембрана соединена с эндоплазматической сетью вокруг пластиды. Вблизи ядра – аппарат Гольджи. В клетке один двухлопастный хроматофор. Он окружен двойной мембраной и хлоропластной эндоплазматической сетью, содержит трехтилакоидные ламеллы. Имеется одна опоясывающая ламелла, в одной из лопастей хлоропласта находится пиреноид, который не удается наблюдать в световом микроскопе. Треть или половину заднего конца клетки занимает крупный пузырь с хризоламинарином. В цитоплазме содержатся еще жир, сократительные вакуоли, митохондрии, микротрубочки, слизевые тельца, которые располагаются непосредственно под цитоплазматической мембраной и состоят из темноокрашенной сердцевины, заключенной в мембрану. Предполагают, что они ответственны за слипание чешуек у М. eaudata.
Клетка покрыта кремнеземными чешуйками, расположенными по спирали. Каждая чешуйка отчасти окружена загнутым краем, отсутствующим в передней части, с многочисленными мелкими порами, почти в центре чешуйки – продолговатое отверстие. Щетинка имеет вид полой трубки с расщелиной на верхушке и несколькими зубцами по одному краю. Своим изогнутым основанием щетинка укреплена в дистальной части чешуйки, где отсутствуют поры. Место образования чешуек и щетинок – пространство между наружной поверхностью хлоропласта и плазмалеммой. Они формируются внутри плоских пузырьков – везикул, которые отпочковываются от хлоропластной эндоплазматической сети и сливаются с везикулами, являющимися дериватами аппарата Гольджи. Концы плоских везикул, производящих чешуйки, вздуты. Примордии и чешуек и щетинок представляют собой плоские пластины. На более поздних стадиях плоские пузыри с пластинами скручиваются в разной степени: в случае щетинок закручивание сильнее. Во время закручивания к везикулам прикрепляются трубчатые элементы, возможно принимающие участие в механизме закручивания. Следствием этого процесса является то, что в трубчатой части щетинки остается щель вдоль одной ее стороны. Клетка продуцирует или чешуйки, или щетинки, но не обе структуры одновременно. Мембрана везикула, содержащего зрелую чешуйку, сливается с плазмалеммой и чешуйка выталкивается в любом месте клеточной поверхности под существующим слоем чешуек. Возможно, чешуйки фиксируются в определенном положении слизью, которая вырабатывается и извергается наружу тельцами, расположенными непосредственно под плазмалеммой. Каким образом чешуйки выстраиваются в правильном порядке по поверхности клетки, пока не ясно. Освобождение щетинок происходит также посредством экзоцитоза. Первой выталкивается из клетки верхушка щетинки; у более длинных щетинок верхушка должна быть вытолкнута из клетки еще тогда, когда ее базальная часть находится в процессе формирования.
У малломонас описан и половой процесс – гологамия, при этом клетки сливаются задними концами.
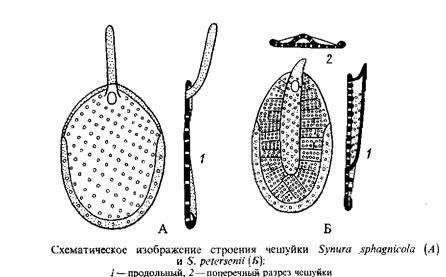
Род синура(Synura) характеризуется шаровидными или эллипсоидальными колониями из грушевидных клеток, задними, иногда очень длинными концами соединяющимися в центре колонии. Кнаружи от цитоплазматической мембраны клетки одеты спирально расположенными, черепитчато перекрывающими друг друга окремнелыми чешуйками, форма и ультраструктура которых, как и в случае малломонас, имеют важное таксономическое значение. Например, у S. sphagnicolaбазальная пластинка на поперечном срезе имеет одинаковую толщину, т.е. плоская. Она пронизана мелкими перфорациями. Передний край снабжен утолщенной апикальной кромкой, базальный край загнут, окружая базальную пластинку наподобие скобы. Полый отогнутый наружу шип прикреплен несколько отступя от переднего края пластинки, у основания его находится пора. Чешуйки других видов, например S. peterscnii, сложнее. Поверх базальной мелко перфорированной пластинки располагается полый медиальный гребень – апикальный, заостренный или тупой, конец которого может выступать за фронтальный край чешуйки, имитируя шип. В передней части медиального гребня находится крупная пора. Базальный конец чешуйки подковообразно загнут и нависает над телом чешуйки. Передние и задние чешуйки, покрывающие тело клетки, несут поперечные ребра, отходящие от медиального гребня, срединные чешуйки, помимо поперечных, имеют еще продольные ребра. Чешуйка не лежит на клетке плоско, а по-видимому, прикреплена только концом, противоположным шипу. Профили чешуек тела у S. sphagnicolaможно наблюдать в цитоплазматических везикулах, большей частью расположенных вблизи наружной поверхности хлоропласта или между хлоропластом и везикулами с хризоламинарином.
Из углубления на переднем конце клетки отходят два гетероконтных и гетероморфных жгутика. Более длинный жгут – с мастигонемами, которые состоят из трубчатой части, несущей терминаль ные нити; короткий жгут – гладкий, с латеральным вздутием. На мембране жгута, как длинного, с мастигонемами, так и короткого, гладкого, располагаются чешуйки жгутиков, размеры и форма которых специфичны для каждого вида и наряду с чешуйками тела клетки служат важным таксономическим признаком. Так, у S. peterseniiчешуйки обоих жгутиков кольцевидные, у S. sphagnicolaна обоих жгутиках чешуйки палочковидные, чешуйки полукруглые, подковообразные, у S. echinulataна длинном жгутике наблюдается смесь булавовидных и кольцевидных чешуек, а на коротком гладком жгуте – только кольцевидные чешуйки. Прикрепляются чешуйки следующим образом: кольцевидные – своими краями, полукруглые – выпуклой стороной, палочковидные – одним концом. В отличие от пропитанных кремнеземом чешуек тела чешуйки жгутиков не минерализованы. Местом образования и сборки чешуек жгутиков у S. sphagnicolaявляется, по-видимому, аппарат Гольджи, у S. peterseniiчешуйки жгутиков наблюдались в пузыре, окруженном расширенной цистерной эндоплазматической сети. Клетка содержит два париетальных хлоропласта, каждый из которых окружен слоем хлоропластной эндоплазматической сети, ламеллы трехтилакоидные, имеется опоясывающая ламелла, глазок отсутствует. Пространство между передними концами хлоропластов занимает одно клеточное ядро, в передней части клетки – аппарат Гольджи, задний узкий конец клетки заполнен сократительными вакуолями, между хлоропластами и поверхностью клетки находятся крупные пузыри, по-видимому, с хризоламинарином.
Клетки внутри колоний делятся продольно. Молодые колонии образуются за счет распада разросшихся старых колоний. Часто одновременно во всех клетках колонии возникают цисты. Известно и пальмеллевидное состояние. У S. peterseniiописан половой процесс – гологамия. Копуляция клеток осуществляется задними концами.
Порядок хризокапсальные – Chrysocapsales
Порядок включает пальмеллоидные формы. Наиболее дифференцированная из пальмеллоидных форм среди водорослей – род гидрурусс единственным видом Н. foeiidus, довольно обычным в быстро текущих холодных горных источниках и представленным оливково-зелеными слизистыми сильно разветвленными тяжами до 30 см длиной, часто издающими очень неприятный запах. Отсюда и видовое название – зловонный. Плотная слизь, образующая цилиндрические тяжи, заключает многочисленные грушевидные клетки с заостренным концом, содержащие один стенкоположный хроматофор, пульсирующие вакуоли и хризоламинарин. Клетки плотно расположены по периферии и более рыхло в центре слизи. На конце каждого тяжа клетка ведет себя как апикальная, одна половинка которой после продольного деления вновь становится апикальной, а другая входит в вещество тяжа. Размножение осуществляется зооспорами характерной тетраэдрической формы. Такая необычная форма зооспор гидруруса определяется сложной системой скелетных микротрубочек, отходящих от жгутикового корня и простирающихся в три передних отростка. Задний отросток также поддерживается сетью микротрубочек. От переднего конца зооспоры отходят два неравной длины жгутика, из которых длинный служит для движения, а более короткий – остаточный – лишен центральных микротрубочек. Зооспоры образуются путем продольного деления клеток более тонких ветвей, клетки более толстых тяжей несут, по-видимому, лишь опорную функцию, не участвуя в размножении. Цисты образуются в некоторых клетках, выступающих из слизистых тяжей,
Класс гаптофициевые, или примнезиофициевые
Этот класс выделен из золотистых водорослей в первую очередь па основе строения монадных клеток, обладающих помимо жгутиков гаптонемой. Включает три порядка, из которых ниже рассматривается один.
Порядок примиезиальные – Prymnesiales
Порядок характеризуется обычно длинной гаптонемой и двумя изоконтными и изоморфными жгутиками. Поверхность клеток кнаружи от плазмалеммы покрыта органическими неминерализованными чешуйками (семейство примнезиевые – Prymnesiaceae) или известковыми телами–кокколитами, в совокупности образующими вокруг клетки коккосферу.
Семейство примнезиевые – Prymnesiaceae. Род хризохромулинапредставлен как в морях, так и в пресных водах. Сферические или овальные клетки с двумя равной длины гладкими жгутиками и гаптонемой кнаружи от цитоплазматической мембраны покрыты органическими неминерализованными чешуйками обычно двух типов, различающимися или только размерами, или же формой.
Так, у Chrysochromulinabirgeriдва типа чешуек, покрывающих тело, различаются только по размерам. Оба они представлены овальными пластинками с рисунком из радиальных гребней, расположенных в квадрантах. Имеются два центральных выступа в виде рогов. У других видов клеточная поверхность покрыта чешуйками, более или менее резко различающимися морфологически. Например, у Ch. cyanophoraвнутренние чешуйки, округлые, плоские, с тонкими концентрическими гребнями, перекрывают друг друга, образуя вокруг клетки футляр, и обычно скрыты под многочисленными наружными цилиндрическими чешуйками.
У Ch. megacyiindraтакже есть два рода чешуек – пластинки и цилиндры. Цилиндры, каждый прикрепленный нижним концом к своей собственной базальной пластинке, распределены по клетке более или менее равномерно, почти касаясь один другого своими боковыми сторонами. Под ними имеется много слоев плоских чешуек с ободками.
'У Ch. chitonнаблюдаются три типа чешуек с характерным расположением: в группах чешуек 6 крупных без ободка располагаются вокруг одной крупной чешуйки с ободком, а все промежутки заполнены самыми мелкими чешуйками.
Семейство кокколитофоридовые – Coccolithophoraceae. К этому семейству относятся почти исключительно морские формы, исключение – пресноводный род гименомонас. Монадные клетки снабжены двумя одинаковыми жгутиками и обычно хорошо заметна гаптонема. Однако у ряда кокколитофорид гаптонема, по-видимому, редуцирована: так, у Н. coronataявной гаптонемы не наблюдалось, у Ochrosphaeraneapolitanaгаптонема представлена только в виде группы микротрубочек, связанных с основаниями жгутиков, и не выступает из клетки. Внутреннее строение клетки такое же, как у других гаптофициевых: ядро, хлоропласты, окруженные хлоропластной эндоплазматической сетью, содержат трехтилакоидные ламеллы, опоясывающая ламелла отсутствует; пиреноид, выступающий от внутренней поверхности хлоропласта и пересеченный парными тилакоидами; аппарат Гольджи, митохондрии и пр. Характерен клеточный покров, расположенный кнаружи от цитоплазматической мембраны и состоящий из пропитанных карбонатом кальция чешуек – кокколитов в совокупности образующих вокруг клетки коккосферу. У ряда форм, помимо кокколитов, имеются неминерализованные органические чешуйки. Тонкая структура кокколитов отличается большим разнообразием и представляет важный таксономический признак. Ниже ограничимся лишь некоторыми примерами кокколитофорид, исследованных в последние годы с помощью электронного микроскопа. Кокколиты, покрывающие клетки Calyptrosphaerapirns, при изучении в сканирующем микроскопе имеют следующее строение: куполообразная верхняя часть отчасти погружена в более широкую, слегка воронковидную базальную часть. На границе между этими двумя частями имеется ложбинка, в которой почти всегда наблюдаются 7–10 перфораций; на куполообразной части кокколита иногда есть углубления. Кокколиты, предположительно окружающие жгутиковый полюс клетки, отличаются от остальных коническим выступом на куполообразной части. У Papposphaeralepidaкокколиты состоят из базальной части с краевым кольцом из пятиугольных элементов; от креста на базальной пластинке отходит вертикальный расширяющийся на конце придаток. У пресноводного рода гименомонас (Hymenomonas) клетки кнаружи от плазмалеммы несут покров из органических чешуек и кокколитов. Последние короновидные, сложенные из 11–12 пятиугольных элементов, расположенных по периферии базальной пластинки.
Изучение некоторых коккофорид в лабораторной культуре позволило обнаружить в цикле их развития неподвижную нитчатую стадию. Например, подвижная одноклеточная стадия Cricosphaeraroscoffensisvar. haptonemoferaможет размножаться путем деления пополам, а при определенных условиях культивирования развивается нитчатая неподвижная стадия, сходная с водорослью Apistonemasubmarlnum.
Как и в случае окремнелых чешуек хризофициевых водорослей, таких, как малломонас, синура и др., неминерализованные органические чешуйки и кокколиты гаптофициевых водорослей в последние десятилетия изучены не только с точки зрения их ультраструктуры, таксономическая значимость которой не вызывает сомнения, но и с точки зрения их генезиса. Из последних исследований в первую очередь заслуживает упоминания работа Р. Броуна с соавторами о синтезе чешуек Pleurochrysisscherffeli.
В 1975 г. были изучены формирование и химический состав чешуек Chrysochromulinachiton. Было показано, что все три типа чешуек разного размера образуются в одно и то же время внутри аппарата Гольджи, затем транспортируются к поверхности клетки, где и располагаются характерным для этого вида образом. То жеотносится к обоим типам чешуек Ch. megacylindra.