Реферат: Отдел золотистые водоросли
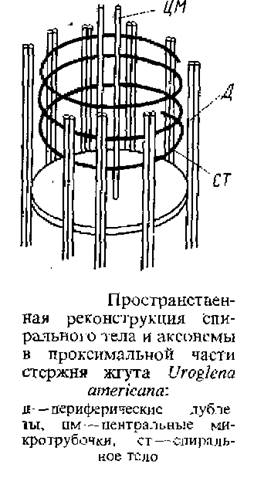
Для водорослей этого отдела характерны хроматофоры, окрашенные в золотистый цвет благодаря тому, что, помимо хлорофиллов а и с, имеется избыток каротиноидов – Р-каротина и ксантофиллов. Для ультраструктуры хлоропластов характерны сложная оболочка, трехтилакоидные ламеллы, локализованный генофор; опоясывающая ламелла у одних имеется, у других отсутствует. Запасные продукты – хризоламинарин, откладывающийся вне хлоропласта и жир. Глазок, заключенный в пластиду, связан со жгутиковым аппаратом. У одних форм жгутики гетероконтные и гетероморфные, у других – изоконтные, изоморфные; у многих одножгутиковых форм электронная микроскопия выявила второй рудиментарный жгутик. Для переходной зоны жгутика характерно спиральное тело, состоящее из 4–5 витков электронно-плотного материала, расположенных между периферическими дублетами и центральными микротрубочками. Золотистые водоросли, стоящие на разных ступенях морфологической дифференцировки талломов, помимо указанных выше признаков, характеризуются особыми эндогенными или эндоплазматическими окремнелыми цистами или статоетюрами, образующимися летом, реже поздней осенью, и служащие для переживания неблагоприятных условий существования. Своеобразие процесса их формирования заключается в том, что оболочка развивается не вокруг протопласта, а закладывается внутри него. Протопласт пространственно разрезается оболочкой на интрацистерную внутреннюю и экстрацистерную наружную части. Экстрацистерная плазма в большей своей части переходит в эндогенно заложившуюся спору, после чего спора закупоривается пробочкой или крышечкой, а Оставшаяся экстрацистерная плазма разрушается. Во многих случаях края поры вытягиваются в виде трубки или расширяются в виде воронки. В то время как оболочка цисты обычно сильно окремневшая, окремнение пробочки значительно меньше, а иногда и отсутствует. При прорастании пробочки растворяются и поделившийся или неподелившийся протопласт споры выходит большей частью в виде монады, снабженной жгутиками, реже в виде безжгутиковой амебы.
Впервые этот процесс эндогенного образования статоспор был описан русским исследователем Л.С. Ценковским в 1870 г. у рода хромулина. Позднее А. Пашер показал, что этот своеобразный способ образования спор свойствен всем золотистым водорослям и встречается не только у монадных и ризоподиальных форм, но и у тех представителей, протопласты которых одеты оболочкой.
В последние годы процесс образования цист у хризомонад, различающихся организацией таллома, был изучен на электронно-микроскопическом уровне. Д. Хибберд исследовал образование цист у голой формы Ochromonastuberculata, К. Сэндгрен у двух видов Dinobryon, у двух колониальных форм – Uroglenavohox, U. americanaи у Mallomonaseaudata–одноклеточной хризомонады, клетка которой имеет покров из окремнелых чешуек. Образование цист происходит в основном сходно у всех изученных до сих пор форм, однако имеются и особенности, которые можно использовать в систематике.
Во всех случаях кремнеземная оболочка цист, как и створки диатомей, образуется внутри пузыря, мембрана которого силиколемма – возникает за счет слияния пузырьков Гольджи. Пузырь в виде сферы, со стенкой, образованной двумя мембранами, в пространстве между которыми и осаждается кремнезем, закладывается внутри цитоплазмы, деля ее на интрацистерную и экстрацистерную.
Когда оболочка цисты сформировалась, внутренняя мембрана пузыря становится функционирующей плазмалеммой цисты; растягиваясь, она перегораживает пору. Наружная мембрана пузыря утрачивается.
Цисты всех хризомонад имеют пору, в зрелом состоянии замкнутую пробкой. Пробка при всем разнообразии структуры у разных видов всегда образуется из материала, освобождающегося из пузырьков Гольджи. которые сливаются с мембраной цисты под порой.
Процесс созревания цист у всех изученных видов сопровождается возрастанием электронной плотности всего содержимого цисты, накоплением липидных гранул. Крупная вакуоль с хризоламинарином приобретает крапчатый вид. Наряду с перечисленными признаками процесса инцистирования, присущими всем хризомонадам, существуют особенности, различающиеся у разных нидон. Некоторые из них, по мнению Сэндгрена, имеют даже филогенетическое значение. Эти особенности касаются: 1) способа развития поры в пузыре, откладывающем кремнезем; 2) отложения кремнезема в пузыре в один или в два этапа в зависимости от того, являются ли оболочки цист гладкими или орнаментированными; 3) судьбы экстрацистерной цитоплазмы и присутствия или отсутствия цитоплазматических микротрубочек, проходящих через пору в экстрацистерную цитоплазму; 4) структуры пробки; 5) числа ядер и пластид в цисте сравнительно с таковым в вегетативных клетках.
1. Пора может возникать двумя путями: или посредством разрыва мембран полностью замкнутого шаровидного пузыря, в котором остается нсокремнелый участок, или же еще во время развития пузыря пора оказывается детерминированной и ее границы очерчены кольцом электронно-плотного' материала. Первый способ образования поры можно наблюдать у Ochromonastuberculata, второй – у Dinobryonи Uroglena.
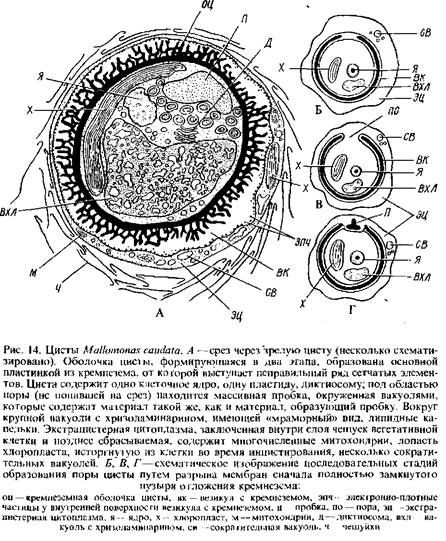
2. У видов с гладкой неорнаментированной оболочкой цисты отмечен лишь один этап и отложении кремнезема внутри пузыря. У форм с орнаментированными цистами после первого этапа отложения внутри пузыря тонкой гладкой окремнелой оболочки наблюдается вторая фаза, когда к основной пластинке из кремнезема снаружи достраиваются шипы иди другая орнаментация, свойственная данному виду. В участках развития орнамтаций пузырь расширяется.
Таким образом, орнаментация оболочек цист, по-видимому, возникает не за счет экстрацистцрной цитоплазмы, как полагал Пашер, а внутри пузыря на втором этапе окремнения. Так, у Dinobryoncylindncumи D. divergensэкстрацистерная цитоплазма, содержащая сократительную вакуоль, втягивается внутрь оболочки цисты до закупорки поры пробкой, и только после этого наступает второй этап окремнения, ответственный за образование шипов на оболочке цист у этих видов. У Mallomonaseaudataкнаружи от стенки цисты остается большое количество цитоплазмы, заключенной в слой чешуек вегетативной клетки. Эта экстрацистерна я цитоплазма с многочисленными митохондриями, пластидой, сократительными вакуолями не втягивается внутрь оболочки цисты до образования пробки, замыкающей пору. Она сбрасывается, когда циста созреет. Оболочка зрелой цисты Mallomonasсостоит из базальной пластинки кремнезема, от которой выступает неправильный ряд сетчатых элементов, возникающих во время второго этапа окремнения. Вдоль внутренней поверхности наружной мембраны пузыря, откладывающего кремнезем, можно видеть плотные гранулы.
3. У изученных видов Dinobryonи Uroglenaэкстрацистерная цитоплазма, содержащая пульсирующую вакуоль, втягивается вну] рь экдоплазматически заложившейся оболочки цисты через пору до образования пробки и таким образом сохраняется. В процессе втягивания экстрацистерной цитоплазмы внутрь оболочки цисты, возможно, играют роль цитоскелетные микротрубочки, выступающие через пору в экс-трацистерную цитоплазму. Напротив, у Ochromonastubercuiataи Maltomonascaudataэкстрацистерная цитоплазма не втягивается внутрь цисты и таким образом не сохраняется: при созревании цисты она сбрасывается.
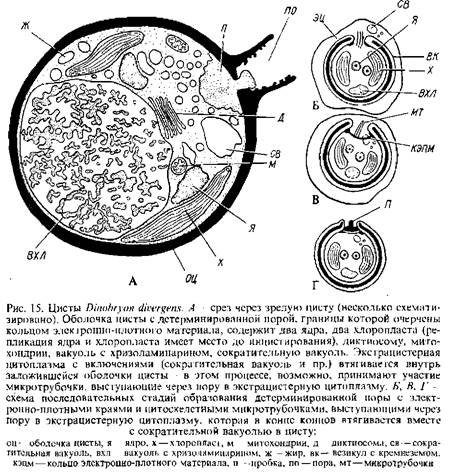
4. Структура пробки, всегда образующейся в результате активности аппарата Гольджи. у разных видов неодинакова. Пробка может быть составлена из разного количества слоев морфологически различающихся материалов, Каждый слой заключает несколько компонентов. Так, пробка у Uroglenaи, возможно, у Mallomonascaudataсодержит только первичный слой. У Ochromonastubercuiataимеется периферический вторичный слой, состоящий из фиброзного материала, заключенного внутри везикул. У Dtnabryoncylindricum, D. divergensпробка цисты также включает второй периферический слой, но состоящий из гранулярного материала. У D. cylindricumимеется еще и третий слой пробки фиброзный. В химическом отношении пробка представляет собой полисахарид и не содержит вообще или содержит лишь небольшие количества кремнезема.
5. Цисты Ochromonastubercuiataи Maltomonascaudataзаключают по одному ядру и одной пластиде, как и вегетативные клетки. Цисты Dinobryonи Uroglenaв отличие от вегетативных клеток содержат по две пластиды и по два ядра. Репликация этих органелл у изученных Сэндреном видов предшествует образованию цист, которые таким образом возникают бесполым путем. Это не исключает возможности появления цист у других видов в результате полового процесса. Такие случаи на уровне оптического микроскопа неоднократно описывались.
При прорастании цисты у Ochromonastubercuiataпробка исчезает и содержимое выходит через пору.
Отдел золотистые водоросли (Chrysophyta) включает невколько классов, из которых ниже приводятся два: класс хризофициевые, или гетерохризофициевые, и сравнительно недавно установленный класс гаптофициевые, или примнезиофициевые, для золотистых водорослей с гаптонемой.
Класс хризофициевые – chrysophyceae
Класс объединяет водоросли с различными типами организации таллома, которые и кладутся в основу деления его на порядки: хризомонадальные, объединяющие мопадные формы, ризохризидальные, представленные золотистыми водорослями с ризоподиальной структурой, хризокапсаль-ные, включающие пальмеллоидные формы, хризосферальные – коккоидные, феотамниальные – нитчатые. Ниже приводятся только два порядка: хризомонадальные и хризокапсальные.
Порядок хризомонадальные нли хромулинальные – Chrysomonadales, Chromulinales
Это самый обширный порядок, включающий водоросли монадной структуры, одноклеточные и колониальные. Систематика хризомонад базируется на числе и строении жгутиков и особенно на характере клеточных покровов. Имеются одной двужгутиковые формы, и прежде считалось, что первые являются исходными, более примитивными. Однако с помощью электронного микроскопа обнаружено, что у кажущихся одножгутиковых форм имеется второй маленький боковой жгутик. Было высказано предположение, что двужгутиковые хризомонады с гетероконтными и гетероморфными жгутиками могли быть исходными, а одножгутиковые формы возникли в результате редукции более короткого жгута. Клеточные покровы хризомонад различны: имеются формы голые, одетые лишь плазмалеммой, у других клетки заключены в целлюлозные домики, у третьих поверх плазмалеммы находится покров из окремнелых чешуек.
Размножение осуществляется делением клеток, у некоторых описан половой процесс.
Хризомонады – преимущественно пресноводные формы, чаще всего приуроченные к чистым водам. Обычно они встречаются в холодное время года, ранней весной и поздней осенью, некоторые – в зимнее время подо льдом. Однако, как было показано для синуры, температура воды здесь имеет лишь косвенное значение. Решающим же фактором является химизм воды, меняющийся на протяжении года: в холодное время года из-за отсутствия другой растительности в воде содержится больше железа и азота. Большинство этих водорослей живут в планктоне, имеют специальные приспособления к планктонному образу жизни и некоторые вызывают «цветение» воды, окрашивая ее в бурый цвет.
Ниже будут рассмотрены представители трех семейств: охромонадовые, или хромулиновые, динобриевые и синуровые.
Семейство Охромонадовые – Ochromonadaceae. Сюда относятся голые формы, клетки которых покрыты только цитоплазматической мембраной с одним или двумя неодинаковыми жгутиками.
Род охромонас, виды которого обычны в планктоне и нейстопе пресных вод, реже солоноватоводных водоемов, представлен одиночными, окрашенными в золотистый цвет клетками с двумя гетероконтными и гетероморфными жгутиками. Клетка охромонас голая, снаружи одетая только цитоплазматической мембраной. Каплевидная форма ее поддерживается цитоскелетом из периферически расположенных микротрубочек. В центре клетки находится клеточное ядро, окруженное ядерной оболочкой из двух мембран. В расширении между мембранами ядерной оболочки заключены два пластинчатых хроматофора с типичной для всего отдела ультраструктурой. В задней части клетки находится крупная вакуоль с хризоламинарииом, в цитоплазме рассеяны митохондрии, в передней части клетки – аппарат Гольджи. От переднего конца клетки отходят два неодинаковых по длине и форме жгутика. Г. Бак проследил тонкое строение и происхождение мастигонем у Ochromonasdanica. Эта водоросль удобна для изучения динамики развития мастигонем, так как ее клетки легко теряют жгутики, а затем образуют их заново. Таким образом можно изучить материал на разных стадиях регенерации жгутикового аппарата. Оказалось, что сами мастигонемы у охромонас сложнее, чем у фукуса и аскофиллума, и в их сборке и выведении на поверхность клетки участвует не только перинуклеарное пространство, но и элементы аппарата Гольджи. Готовые трубчатые мастигонемы охромонас состоят из базальной части, прикрепляющейся к мембране жгута, микротрубчатой части и 1–3 терминальных нитей. Кроме этих компонентов, сходных с таковыми мастигонем бурых водорослей, трубчатый стержень мастигонемы по всей поверхности несет длинные и короткие латеральные нити. Трубчатые мастигонемы располагаются на длинном жгутике в два неодинаковых ряда: с одной стороны мастигонемы прикрепляются к специальным участкам жгутиковой мембраны поодиночке, а на противоположной стороне жгута – пучками по три – пять. Помимо трубчатых мастигонем, длинный жгут несет тонкие простые нитевидные фиброзные мастигонемы. Короткий жгутик имеет только фиброзные мастигонемы. На рис. 16, В представлена схема возможной последовательности сборки мастигонем и их освобождения при регенерации жгутика у охромонас. По мере удлинения жгута новые мастигонемы и, возможно, мембрана, к которой они прикрепляются, добавляются к основанию жгута.
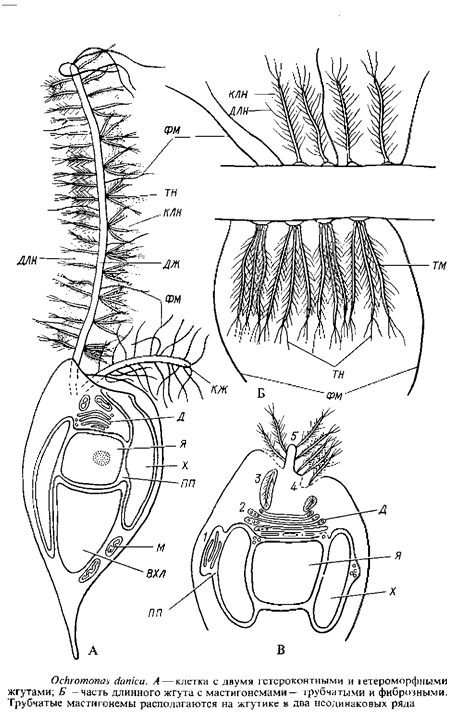
Такое же образование промастигонем и выделение готовых мастигонем поодиночке или в группах, наружу к основанию жгутика в цистернах аппарата Гольджи наблюдалось и у Ochromonasminuta. Жгутиковый корень у охромонаса состоит из поперечно исчерченных пучков фибрилл, которые, разветвляясь, достигают поверхности клеточного ядра.
--> ЧИТАТЬ ПОЛНОСТЬЮ <--