Реферат: Получение газообразного и жидкого топлива. Получение биогаза, метана, спиртов, тепловой энергии
Органические кислоты, спирты,
NH![]() ,СО2 и Н2
,СО2 и Н2
|

![]()
![]() СО2 + СН4
СО2 + СН4
Рис. 1 Деструкция биополимеров до метана природными ассоциациями микроорганизмов, протекающая в 3 стадии
В зависимости от температуры протекания процесса метановые бактерии разделяют на мезо- и термофильные. Оптимальная температура для мезофильных бактерий от 30 до 40°С, а для термофильных от 50 до 60°С. В целом термофильный процесс метаногенеза идет интенсивнее мезофильного, притом в этих условиях анаэробной переработки отходов субстрат обеззараживается от патогенной микрофлоры и гельминтов. При анаэробной переработке отходов животноводческих ферм микрофлора метантенков (анаэробных ферментеров) формируется преимущественно из микрофлоры желудочно-кишечного тракта данного вида животных и микрофлоры окружающей среды. Из наиболее часто встречающихся культур следует отметить Lactobacillus acidophilus, Butyrivibrio fibrisolvens, Peptostreptococcus productus, Bacteroides uniformis, Eubacterium aerofaciens.
К числу целлюлозоразлагающих бактерий микрофлоры жвачных относятся Bacteroides succinoqenes и Ruminococcus flavefaciens. Из рубца и навоза жвачных были изолированы такие метанообразующие бактерии, как Methanobacterium mobile, Methanobrevibacter rumi- nantium и Methanosarcina ssp . После определенного срока работы метантенка при установленном температурном режиме и на постоянном субстрате образуется сравнительно стабильный консорциум микроорганизмов. В ходе изучения микрофлоры свиного навоза при метановом брожении выделено около 130 различных бактерий.
Первую стадию разрушения сложных органических полимеров осуществляют бактерии семейств Enterobacteriaceae, Lactobacillaceae, Streptococcaceae, атакже представители родов Clostridium, Butyrivibrio, Bacteroides, Rumino coccus и некоторых других способных к брожению.Главные продукты ферментации — ацетат, пропионат, сукцинат, Н2 и СО2 . Конечными продуктами ферментации целлюлозы и гемицеллюлозы под действием бактерий, выделенных из рубца жвачных и кишечника свиней, являются различные летучие жирные кислоты.
Бактерии второй, или ацетогенной, фазы, относящиеся к родам Syntrophobacter, Syntrophomonas и Desulfovibrio, вызывают разложение пропионата, бутирата, лактата и пирувата до ацетата, Н2 и СО2 — предшественников метана. Ряд микроорганизмов способны синтезировать ацетат из СО2 в термофильных условиях, к их числу принадлежат Clostridium formicoaceticum, Acetobacterium woodii, метановые бактерии из родов Methanothrix, Methanosarcina, Methanococcus, Methanogenium и Methanospirillum.
Свойства метанобразующих бактерий
Полагают, что метанобразующие бактерии, или метаногены, возникли около 3,0—3,5 млрд. лет тому назад и достигли своего расцвета в архее.
Сейчас эти микроорганизмы имеют достаточно широкое распространение, приуроченное к анаэробным условиям. Вместе с другими бактериями они активно участвуют в деструкции органических веществ в различных экологических нишах: в болотном, речном и озерном иле, в осадках морей и океанов, в искусственных технических сооружениях — метантенках, в рубце жвачных и пищеварительном тракте ряда других животных.
Среди этих бактерий есть организмы, клетки которых близки к сферическим, образующие агрегаты, похожие на сарцин, ланцетовидные, палочковидные и нитевидные формы. Большинство неподвижны, но отдельные виды проявляют способность к движению в результате наличия жгутиков.
Как и другие архебактерии, метаногены отличаются от остальных прокариот (эубактерий) составом ряда компонентов клеток, в том числе клеточной стенкой, не содержащей муреина, а также характером липидов, в которые не входят жирные кислоты. Большую часть нейтральных липидов составляют простые эфиры глицерина и длинноцепочечного спирта фитанола. В метаболизме метанобразующих бактерий участвует кофермент М (2-меркаптоэтансульфоновая кислота), факторF420 , представляющий собой особый флавин, и ряд других соединений, которые у остальных организмов не обнаружены или встречаются крайне редко. Но главное, что отличает метаногенов, как и других архебактерий, от остальных организмов, — это нуклеотидная последовательность в 16S рРНК, о чем свидетельствует изучение состава получаемых из нее олигонуклеотидов.
В то же время отдельные виды метанобразующих бактерий по этому показателю, а также содержанию ГЦ в ДНК (от 30,7 до 61,0 мол. %) могут отличаться весьма значительно. На этом основании их подразделяют на три порядка, включающих несколько семейств и родов. Наиболее изученными видами являются Methanobacterium thermoautotrophicum, Methanosarcina barkeri, Methanobrevibacter ruminantium.
Все метанобразующие бактерии — строгие анаэробы. Как уже говорилось выше, они делятся на мезо- и термофилов. К числу термофилов относится, например, М. thermoautotrophicum. Оптимальное значение рН для роста разных видов 6,5—8,0. Некоторые штаммы способны расти при наличии в среде до 5—7 % и более NaCl.
Как источник серы бактерии чаще всего используют сульфид, а как источник азота — аммоний. Некоторые виды нуждаются для роста в наличии дрожжевого автолизата или смеси витаминов. Известны также метанобразующие бактерии, для роста которых необходимо присутствие ацетата и (или) других органических веществ.
Но довольно многие из этих микроорганизмов могут расти в автотрофных условиях (при наличии в качестве единственного источника углерода углекислоты).
Ассимиляция углекислоты в автотрофных условиях, как показано для М. thermoautotrophicum, происходит особым нециклическим путем. Началом его является образование из углекислоты С2 -соединения в виде, ацетил-кофермента А (КоА). Дальнейшее превращение ацетил-КоА включает следующие реакции:
ацетил-КоА![]() пируват
пируват![]() фосфоенолпируват
фосфоенолпируват![]()
![]() оксалоацетат
оксалоацетат![]() малат
малат![]() фумарат
фумарат![]() сукцинат
сукцинат![]()
![]() сукцинил-КоА
сукцинил-КоА![]() α-кетоглутарат.
α-кетоглутарат.
Образующиеся пируват, оксалоацетат, 2-кетоглутарат используются частично для синтеза аминокислот, а фосфоенолпируват — углеводов.
Субстратами, из которых большинство известных видов метаногенов образуют метан, служат молекулярный водород и углекислота:
4Н2 + СО2 ![]() СН4 + 2Н2 О
СН4 + 2Н2 О
Ряд видов синтезируют метан из окиси углерода:
4СО + 2Н2 О ![]() СН4 + ЗСО2
СН4 + ЗСО2
Некоторые виды образуют метан из муравьиной кислоты:
4НСООН ![]() СН4 + ЗСО2 + 2Н2 О
СН4 + ЗСО2 + 2Н2 О
Известны метанобразующие бактерии, образующие метан из метанола, метиламина, диметиламина, триметиламина и (или) ацетата:
4СН3 ОН![]() ЗСН4 + СО2 + 2Н2 О
ЗСН4 + СО2 + 2Н2 О
4CH3 NH2 + 2Н2 О![]() ЗСН4 + CO2 + 4NH3
ЗСН4 + CO2 + 4NH3
СНзСООН![]() СН4 + СО2
СН4 + СО2
При превращении метилированных аминов, метанола и ацетата в метан в него переходит метильная группа субстрата.
Еще в 1956 г. известным биохимиком И. X. Баркером предложена схема восстановления СО2 в метан, которая сейчас детализирована (рис. 2). Эта схема послужила отправной точкой для формулирования гипотезы о том, что метанобразующие бактерии могут аккумулировать энергию в процессе окислительного фосфорилирования.
В исследованиях, проведенных с чистыми культурами разных видов метаногенов, установлено, что синтез метана протекает в мембранах, причем этот процесс сопряжен с генерацией трансмембранного потенциала, энергия которого трансформируется в АТФ.
Таким образом, метанобразующие бактерии, видимо, осуществляют (во всяком случае, когда окисляют Н2 ) не брожение, а анаэробное дыхание, используя СО2 в качестве акцептора электронов. Но выход энергии на каждый образованный моль метана у метаногенов не превышает двух молей АТФ. Поэтому для обеспечения своего роста метанобразующие бактерии должны синтезировать значительные количества метана. Из 100 % метаболизированного соединения углерода они трансформируют в клеточный материал только 5-10 %, остальное конвертируется в метан.
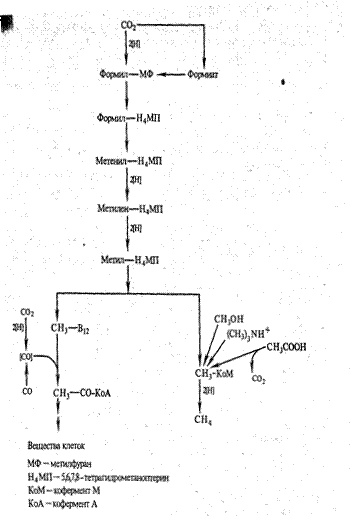
Рис. 2. Схема образования метана из СО2 и других соединений