Реферат: Популяции
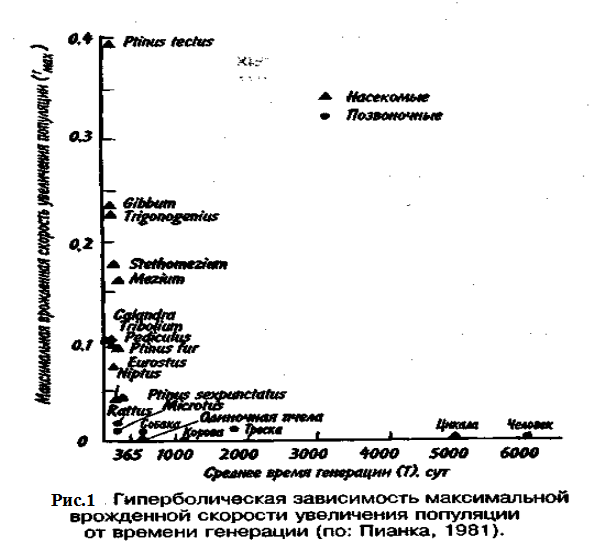
Тем не менее преимущества, даваемые большими размерами тела, должны превышать недостатки, связанные с уменьшением rтах , так как в противном случае крупные организмы никогда не появились бы в эволюции. Тенденция к возрастанию размеров тела с течением геологического времени, прослеженная по ископаемым остаткам, послужила основанием для введения понятия увеличение филетического размера.
Крупные размеры тела дают совершенно очевидные преимущества: более крупный организм должен привлекать меньше потенциальных хищников и, следовательно, он имеет больше шансов не стать жертвой и должен отличаться лучшей выживаемостью; мелкие организмы находятся в тесной зависимости от физической среды, даже очень слабые изменения которой могут оказаться для них губительными. Более крупные организмы легче переносят воздействие подобных изменений и соответственно лучше от них защищаются. Однако более крупным организмам требуется больше вещества и энергии в расчете на одну особь в единицу времени, чем мелким. Кроме того, для них существует гораздо меньше укрытий и безопасных мест.
В жизни всех организмов в популяции можно выделить три основных периода: предрепродуционный, репродукционный и пострепродукционный. Относительная продолжительность каждого из них у разных видов весьма различна. У многих животных первый период самый длинный. Ярким примером могут служить поденки, у которых предрепродуционный период достигает 3 лет, а репродукционный занимает всего от 2-3 ч до 1 суток. У американской цикады предрепродуционный период составляет 17 лет. Но есть виды, особи которых, едва появившись на свет, начинают интенсивно размножаться (большинство бактерий).
Репродукционные возможности популяции зависят от ее возрастного состава. Продолжительность жизни особей популяции можно оценить, используя кривые выживания. Существует три типа кривых выживания (рис. 2).
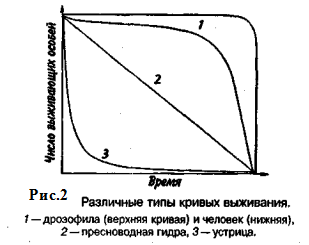
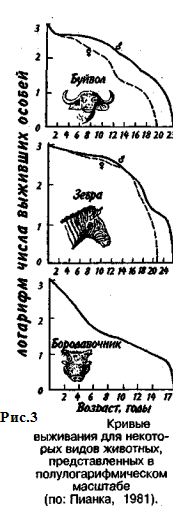
Первый тип (кривые 1) соответствует ситуации, когда большее число особей имеет одинаковую продолжительность жизни и умирает в течение очень короткого отрезка времени. Кривые характеризуются сильно выпуклой формой. Такие кривые выживания свойственны человеку (рис. 2, 1), причем кривая выживания для мужчин по сравнению с аналогичной кривой для женщин менее выпуклая, поэтому страховой полис для мужчин в большинстве стран Запада в 1,5 раза дороже, чем для женщин. Для большинства копытных кривые выживания также выпуклые (рис. 3), хотя и в различной степени для разных видов, а также в зависимости от пола. Второй тип (рис. 2, 2) свойствен видам, коэффициент смертности которых остается постоянным на протяжении всей их жизни. Поэтому кривая выживания трансформируется в прямую линию. Такая форма кривой выживания свойственна пресноводной гидре. Третий тип (рис. 2, 3) — сильно вогнутые кривые, отражающие высокую смертность особей в раннем возрасте. Так характеризуется продолжительность жизни некоторых птиц, рыб, а также многих беспозвоночных.
Знание типа кривой выживания дает возможность построить пирамиду возрастов (рис. 4). Следует различать три типа таких пирамид. Пирамида с широким основанием, что соответствует высокому проценту молодняка, характерна для популяции с большим значением коэффициента рождаемости. Средний тип пирамиды соответствует равномерному распределению особей по возрастам в популяции со сбалансированными коэффициентами рождаемости и смертности — пирамида выровненная. Пирамида с узким основанием (обращенная), отвечающая популяции с численным преобладанием старых особей над молодняком, характерна для сокращающихся популяций. В таких популяциях коэффициент смертности превышает коэффициент рождаемости.


Большое значение для увеличения численности популяции имеют затраты на потомство, выражающиеся в определенной тактике размножения. Не все потомки равноценны: те из них, которые произведены на свет в конце вегетационного сезона, обычно имеют меньшую вероятность дожить до взрослого состояния по сравнению с потомками, появившимися на свет раньше. Сколько усилий должны родители затрачивать на каждого потомка? При постоянной величине репродуктивного усилия средняя приспособленность отдельного потомка связана обратным соотношением с их числом. Один крайний вариант тактики размножения — вложить все в единственного очень крупного и хорошо приспособленного потомка, другой — максимально увеличить общее число произведенных на свет потомков, вложив в каждую особь как можно меньше. Однако наилучшая тактика размножения —это компромисс между производством максимально большого числа потомков и образованием потомства максимально высокой приспособленности.
Указанное соотношение количества и качества потомства иллюстрирует простая графическая модель (рис. 5).
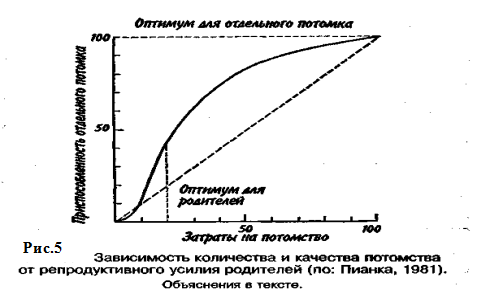
В маловероятном случае, т. е. в случае линейной зависимости приспособленности потомков от затрат на них родителей, приспособленность каждого отдельного потомка уменьшается с увеличением размера помета или кладки. Так как приспособленность родителей или, что то же самое, общая приспособленность всех потомков - постоянная величина, с точки зрения родителей не существует оптимального размера кладки. Однако поскольку первоначальные расходы на потомство вносят больший вклад в приспособленность потомков, чем последующие (имеет место 5-образный характер зависимости приспособленности потомков при увеличении вклада родителей; см. рис. 7.6), то очевидно, что существует некоторый оптимальный размер кладки. В данном гипотетическом случае родители, расходующие только 20% своего репродуктивного усилия на каждого из пяти своих потомков, получат большую отдачу от своего вклада, чем при любом другом размере кладки. Подобная тактика, будучи оптимальной для родителей, не является наилучшей для каждого отдельно взятого потомка, максимальная приспособленность которого достигается в том случае, если он единственный отпрыск, получивший полный вклад усилий от своих родителей. Следовательно, в данном случае налицо «конфликт родителей и детей».
Особенно большое влияние на форму S-образной кривой оказывает конкурентная обстановка. В сильно разреженной среде (конкурентный вакуум) наилучшей репродуктивной стратегией следует считать максимальный вклад вещества и энергии в размножение для продуцирования как можно большего количества потомков в самые короткие сроки. Поскольку конкуренция невелика, потомки могут выжить, даже если они имеют очень малые размеры и низкую приспособленность. Однако в насыщенной среде обитания, где заметно проявляются эффекты массы, а конкуренция остра, оптимальной стратегией будет расходование большого количества энергии на преодоление конкуренции, повышение собственной выживаемости и на продуцирование более конкурентоспособных потомков. При подобной стратегии лучше иметь крупных потомков, а поскольку энергетически они дороже, их может быть произведено на свет меньше.
Итак, свойства популяции можно оценить по таким показателям, как рождаемость, смертность, возрастная структура, соотношение полов, частота генов, генетическое разнообразие, скорость и форма кривой роста и т. д.
Плотность популяции определяется ее внутренними свойствами, а также зависит от факторов, действующих на популяцию извне.
ФАКТОРЫ ДИНАМИКИ ЧИСЛЕННОСТИ ПОПУЛЯЦИЙ
Известно три типа зависимости численности популяции от ее плотности (рис. 6). При первом типе (кривая 1) скорость роста популяции уменьшается по мере увеличения плотности. Это широко распространенное явление позволяет понять, почему популяции некоторых животных относительно устойчивы. Во-первых, при увеличении плотности популяции наблюдается снижение рождаемости. Так, в популяции большой синицы при плотности меньше одной пары на 1 га на одно гнездо приходится 14 птенцов; когда же плотность достигает 18 пар на 1 га, выводок составляет менее 8 птенцов. Во-вторых, при увеличении плотности популяции меняется возраст наступления половой зрелости. Например, африканский слон в зависимости от плотности популяции может достигать половой зрелости в возрасте от 12 до 18 лет. Кроме того, этот вид при низкой плотности дает приплод 1 слоненок за 4 года, тогда как при высокой — рождаемость составляет 1 слоненок за 7 лет.
При втором типе зависимости (кривая 2) темп роста популяции максимален при средних, а не при низких значениях плотности. Так, у некоторых видов птиц (например, чаек) число птенцов в выводке увеличивается с повышением плотности популяции, а затем, достигнув наибольшей величины, начинает уменьшаться. Этот тип влияния плотности популяции на скорость размножения особей характерен для видов, у которых отмечен групповой эффект. При третьем типе (кривая 3) темп роста популяции не изменяется до тех пор, пока она не достигнет высокой плотности, затем резко падает.
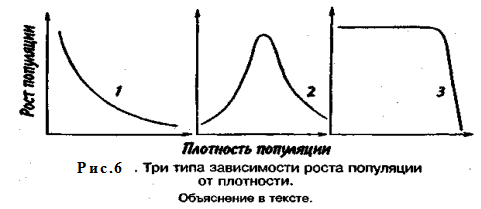
Подобная картина наблюдается, например, у леммингов. При пике численности плотность леммингов становится избыточной, и они начинают мигрировать. Элтон так описал миграции леммингов в Норвегии: животные проходили через деревни в таком количестве, что собаки и кошки, которые вначале нападали на них, просто перестали их замечать. Достигнув моря, обессиленные лемминги тонули.
Регуляция численности равновесных популяций определяется преимущественно биотическими факторами. Среди них главным фактором часто оказывается внутривидовая конкуренция. Примером может служить борьба птиц за мести для гнездования.
Внутривидовая конкуренция может быть причиной физиологического эффекта, известного под названием шоковой болезни. Его отмечают, в частности, у грызунов. Когда плотность популяции становится слишком большой, шоковая болезнь приводит к снижению плодовитости и увеличению смертности, что возвращает плотность популяции к нормальному уровню.
У некоторых видов животных взрослые особи питаются собственным приплодом. Это явление, известное как каннибализм, снижает численность популяции. Каннибализм свойствен, например, окуням: в озерах Западной Сибири 80% пищи крупных особей составляет молодь того же вида. Молодь, в свою очередь, питается планктоном. Таким образом, когда нет других видов рыб, взрослые особи живут за счет планктона.
Межвидовые взаимодействия также играют существенную роль в контроле плотности популяции. Взаимодействия паразит-хозяин и хищник-жертва часто за висят от плотности. Болезни также являются фактором, принимающим участие в регуляции плотности популяции. Когда кролики болеют миксоматозом, вызываемым вирусом, распространение инфекции идет гораздо быстрее в популяциях с повышенной плотностью.
Хищничество как ограничивающий фактор само по себе имеет большое значение. Причем если влияние жертвы на численность популяции хищника не вызывает сомнений, то обратное воздействие, т. е. на популяцию жертвы, бывает не всегда. Во-первых, хищник уничтожает больных животных, тем самым он улучшает средний качественный состав популяции жертвы. Во-вторых, роль хищника ощутима только тогда, когда оба вида обладают приблизительно одинаковым биотическим потенциалом. В противном случае из-за низкого темпа размножения хищник не в состоянии ограничить численность своей жертвы. Например, только одни насекомоядные птицы не могут остановить массовое размножение насекомых. Иными словами, если биотический потенциал хищника намного ниже биотического потенциала жертвы, действие хищника приобретает постоянный характер, не зависящий от плотности его популяции.
Численность насекомых-фитофагов нередко определяется комбинацией видоспецифичных реакций насекомых и растений на воздействие загрязняющих веществ. Загрязнение понижает резистентность растений, в результате чего численность насекомых возрастает. Однако при слишком большом загрязнении численность насекомых падает, несмотря на уменьшение сопротивляемости растений.
Приведенная дифференциация факторов динамики численности популяций позволяет понять их реальное значение в жизни и воспроизводстве популяций. Современная концепция автоматического регулирования численности популяций базируется на сочетании двух принципиально различных явлений: модификаций, или случайных колебаний численности, и регуляций, действующих по принципу кибернетической обратной связи и нивелирующих колебания. В соответствии с этим выделяют модифицирующие (независящие от плотности популяции) и регулирующие (зависящие от плотности популяции) экологические факторы, причем первые из них воздействуют на организмы либо непосредственно, либо через изменения других компонентов биоценоза. По существу, модифицирующие факторы представляют собой различные абиотические факторы. Регулирующие факторы связаны с существованием и активностью живых организмов (биотические факторы), поскольку лишь живые существа способны реагировать на плотность своей популяции и популяций других видов по принципу отрицательной обратной связи (рис. 7).