Реферат: Происхождение хордовых животных
Хордовые - один из крупнейших типов животного царства, представители которого освоили все среды обитания. В состав этого типа входят три группы (подтипа) организмов: оболочники (в том числе живущие на дне морские сидячие организмы - асцидии), бесчерепные (похожие на маленьких рыбок морские существа - ланцетники), позвоночные (хрящевые и костистые рыбы, земноводные, пресмыкающиеся, птицы и млекопитающие). Человек - тоже представитель типа хордовых. Происхождение типа хордовых - это важнейший этап в историческом развитии животного мира, означающий появление группы животных с уникальным планом строения, который позволил в дальнейшей эволюции достичь максимальной среди живых существ сложности строения и поведения. В сущности это означает реконструкцию первых шагов на том пути, который привел к человеку. Вот почему проблема происхождения хордовых уже более полутора столетий вызывает большой интерес биологов.
Происхождение хордовых относится к тому периоду эволюции животного царства, от которого сохранилось очень мало палеонтологических остатков. Известно лишь, что в среднем кембрии (около 550 млн. лет назад) уже существовали бесчерепные, родственные современным ланцетникам. Более ранние этапы эволюции хордовых приходится реконструировать на основе изучения современных организмов, преимущественно сравнительно-анатомическими и эмбриологическими методами, привлекая данные и из других областей биологии.
Основные гипотезы происхождения хордовых
Одна из первых концепций, связывающих план строения хордовых с планами строения других групп беспозвоночных, была разработана выдающимся французским сравнительным анатомом Жоффруа Сент-Илером еще в первой половине XIX века. Он считал, что членистые животные (кольчатые черви и членистоногие) могут рассматриваться как перевернутые позвоночные. "Взгляните на рака, опрокинутого на спину, говорил он, - и вы увидите, что различные системы его расположены совершенно так же, как у высших позвоночных животных". Именно эта концепция была в центре публичного спора между Жоффруа Сент-Илером и Жоржем Кювье, который состоялся в Париже в 1830 году в дни июльской революции, свергнувшей Бурбонов. Как известно, в том знаменитом споре победил Жорж Кювье. Однако к идее сравнения хордовых (и позвоночных) и перевернутых кольчатых червей или членистоногих зоологи возвращались неоднократно на протяжении всего XIX века.
Эту точку зрения отстаивал выдающийся зоолог и, как бы мы сейчас сказали, "организатор науки" Антон Дорн, построивший на свои личные средства первую в мире и до сих пор успешно работающую Неаполитанскую зоологическую станцию. В начале XX века идею происхождения хордовых от перевернувшихся на спинную сторону членистоногих отстаивали Гаскелл, Паттен и другие биологи.
Остроумные и хорошо объясняющие план строения хордовых гипотезы их происхождения от перевернутых аннелид или членистоногих утратили свою популярность прежде всего в связи с тем, что стали ясны фундаментальные эмбриологические отличия между кольчатыми червями и членистоногими, с одной стороны, и хордовыми - с другой.
В начале нынешнего столетия были разработаны гипотезы происхождения хордовых от кишечнодышащих - обитающих в толще грунта морских червеобразных организмов. У этих животных (как и у хордовых) имеются жаберные щели, а в передней части тела - опорный вырост кишечника (стомохорд), который сравнивали с хордой. Однако кишечнодышащие резко отличались от хордовых обратным направлением тока крови, расположением сердца, строением нервной системы и другими важными особенностями организации. Делались попытки выводить хордовых от иглокожих, немертин, моллюсков и других групп беспозвоночных, однако ни одна из них не имела успеха.
Своеобразной реакцией на неудачи зоологов решить проблему происхождения хордовых традиционными способами явилась разработка так называемых неотенических гипотез, выводящих хордовых от личинок асцидий. Взрослые асцидии - прикрепленные морские животные, фильтрующие воду через объемистый жаберный мешок и улавливающие взвешенные в воде мельчайшие органические частицы. Они не имеют ни хорды, ни нервной трубки и совсем не похожи на хордовых животных. Но, как давно известно (а впервые это показал еще в конце прошлого века выдающийся русский биолог А.О. Ковалевский), личинки асцидий имеют и хорду, и нервную трубку, которые утрачиваются при метаморфозе. На основании этого многие биологи (Берилл, Уитер, Боун и др.) предполагают, что хордовые произошли от ставших половозрелыми личинок асцидий. Правда, в этом случае остается непонятным, откуда взялись сами асцидии. Да и трудно представить, чтобы хорда и нервная трубка возникли у живущих 1 - 2 дня и не питающихся личинок. Более вероятно, что эти особенности - наследие свободноживущих предков асцидий, об организации которых неотенические гипотезы ничего не говорят. Ни одна из рассмотренных гипотез в настоящее время не может считаться истинной, но в каждой из них содержатся отдельные элементы, которые могут оказаться полезными при реконструкции путей происхождения хордовых.
Первичноротые и вторичноротые
Все билатерально-симметричные организмы делятся на две группы - первичноротые и вторичноротые, и раньше чем приступить к анализу проблемы происхождения хордовых, следует выяснить, к какой из этих двух групп принадлежат хордовые. Названия "первичноротые" (Protostomia) и "вторичноротые" (Deuterostomia) произошли от способа развития ротового отверстия в эмбриогенезе. Так, у первичноротых животных бластопор (возникающее в эмбриональном развитии отверстие первичного кишечника) частично или полностью переходит в ротовое отверстие. У вторичноротых (например, у иглокожих) бластопор становится анальным отверстием, а рот прорывается заново на переднем конце личинки. В других случаях бластопор закрывается, а рот и анус открываются заново.
Помимо судьбы рта, первичноротые и вторичноротые отличаются другими эмбриологическими особенностями. Во-первых, это характер дробления оплодотворенного яйца: у первичноротых оно спиральное, а у вторичноротых - радиальное (рис. 1). Другое отличие состоит в способе закладке целома (вторичной полости тела): у первичноротых стенки вторичной полости тела происходят от двух клеток, а у вторичноротых за счет выпячивания карманов эмбрионального кишечника (рис. 1). Третьим важнейшим различием между первично- и вторичноротыми является судьба зачатка первичного мозга: у первичноротых он дает начало мозгу взрослых форм, а у вторичноротых всегда редуцируется, так что новый нервный центр возникает в другом месте заново. Вторичноротые - это вторичномозговые животные.
Рот и анус хордовых животных формируются без связи с бластопором. Хордовые имеют типичное радиальное дробление яйца, вторичная полость тела (целом) формируется у них за счет выпячивания карманов первичного кишечника, а в нервной системе нет следов первичного мозга. Все эти признаки указывают на принадлежность хордовых к вторичноротым животным. Помимо хордовых к ним относятся погонофоры, полухордовые и иглокожие (рис. 2). В состав первичноротых входят плоские и круглые черви, кольчатые черви, моллюски, членистоногие (рис. 2).
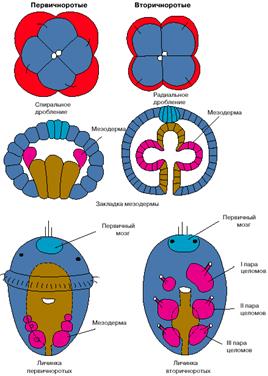
Рис. 1. Особенности эмбрионального развития первичноротых и вторичноротых животных.
Сравнение хордовых с другими вторичноротыми
Хотя хордовые - это вторичноротые животные, все же их развитие имеет ряд особенностей. На рис. 3 представлены соответствующие стадии развития одной из групп вторичноротых - погонофор и низших хордовых - асцидий.
Анимальный полюс яйца погонофор, на котором располагается ядро яйцеклетки, соответствует спинной стороне эмбриона, а вегетативный, на котором концентрируется желток, - брюшной. У низших хордовый ситуация обратная: анимальный полюс соответствует брюшной стороне, а вегетативный - спинной. Бластопор (то есть та область эмбриона, где впячивается энтодерма и формируется первичный кишечник) у погонофор соответствует брюшной стороне личинки, а у низших хордовых - спинной. Зачаток нервной системы, закладывающийся по линии замыкания бластопора, у погонофор становится брюшным нервным тяжем, а у хордовых - спинной нервной трубкой (рис. 3).
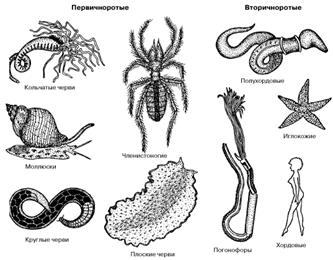
Рис. 2. Представители основных групп первичноротых и вторичноротых животных.
План строения низших хордовых также существенно отличается от планов строения других вторичноротых. Так, например, у всех вторичноротых передние целомы диссимметричны: левый передний целом больше правого, который обычно выполняет функцию околосердечной сумки. У хордовых ситуация обратная: правый передний целом больше левого. Целомы вторичноротых сообщаются с внешней средой отверстиями, открывающимися на спинной стороне и выполняющими осморегуляторные и половые функции. У хордовых эти отверстия располагаются на той стороне тела, которую мы традиционно считаем брюшной.
Кровеносная система у вторичноротых и хордовых устроена по общему плану и состоит из двух главных сосудов: спинного и брюшного. У вторичноротых, как и всех других беспозвоночных, кровь по спинному сосуду течет вперед, а по брюшному назад. У хордовых направление тока крови обратное: по спинному сосуду кровь течет назад, а по брюшному - вперед. Сердце у вторичноротых лежит на спинной стороне, а у хордовых - на брюшной.
Суммируем все эти отличия.
1. Бластопор хордовых соответствует не брюшной, а спинной стороне.
2. Нервная пластинка хордовых, закладывающаяся по месту замыкания бластопора, превращается не в брюшной нервный тяж, а в спинную нервную трубку.
3. Диссимметрия первой пары целомов имеет обратный знак в сравнении с другими вторичноротыми: правый передний целом у хордовых больше левого.
4. Отверстия целомов у хордовых открываются не на спинной, а на брюшной стороне.
5. Кровь у хордовых по брюшной стороне течет вперед, а по спинной - назад, и сердце располагается не на спинной, а на брюшной стороне.
Что же означают эти несоответствия? Только одно: предки хордовых в своем эволюционном развитии испытали инверсию сторон тела, то есть перевернулись и стали передвигаться на морфологически спинной стороне, которая стала функционировать как физиологически брюшная. Морфологически брюшная сторона предков хордовых стала функционировать как физиологически спинная.
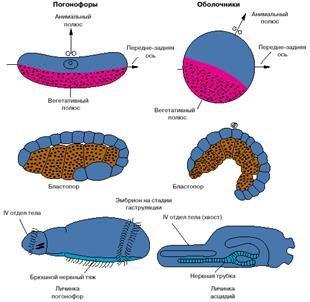
Рис. 3. Особенности эмбрионального развития погонофор и низших хордовых.
Если мы примем этот вывод, все перечисленные выше несоответствия исчезнут: бластопор хордовых станет соответствовать их исходно брюшной стороне, нервная трубка будет располагаться на морфологически брюшной стороне, диссимметрия передних целомов приобретет нормальный для вторичноротых характер, отверстия целомов окажутся на морфологически спинной стороне, и направление тока крови, так же как положение сердца, окажется типичным для вторичноротых. На рис. 4 представители низших хордовых (личинка асцидий и ланцетник) изображены перевернутыми, чтобы продемонстрировать, что при таком положении их системы органов соответствуют таковым других вторичноротых.
Переворот на спинную сторону мог стать тем важнейшим эволюционным событием, которое определило формирование плана строения хордовых животных и его коренное отличие от планов строения вторичноротых и других беспозвоночных.
--> ЧИТАТЬ ПОЛНОСТЬЮ <--