Реферат: Психрофільні мікроорганізми та їх використання
2.1 Вплив температури на ріст та розмноження
Ріст – основна функція мікроорганізмів, на яку впливає температура. Ріст підпорядковується термодинамічним законам, хоча має і свої специфічні особливості.
Основною характеристикою темпу розмноження мікроорганізмів є питома швидкість росту. Середня питома швидкість росту за проміжок часу (t1 -t2 ) розраховується за формулою 1:
![]() де: m1 таm0 – початкова і кінцева вага клітин.
де: m1 таm0 – початкова і кінцева вага клітин.
Формула 1 Швидкість росту
Температура, при якій швидкість росту максимальна називається оптимальною.
Головною особливістю психрофілів, на відміну від інших мікроорганізмів, є здатність інтенсивно рости при температурах 0–10 °С. Прикладом можуть послугувати такі вибіркові данні:
Табл. 1. Час генерації різних штамів Pseudomonas при температурах від 26–0 °С
|
Температура в °С | Час генерації в хв. | |
|
Pseudomonas 1–36 (психрофіл) | Pseudomonas aeruginosa (мезофіл) | |
| 26 | 59 | - |
| 25 | - | 56 |
| 20 | 95 | 120 |
| 15 | - | 160 |
| 14 | 140 | - |
| 10 | 160 | 470 |
| 8 | 210 | 1400 |
| 0 | 620 | - |
Можна також привести данні про інтенсивність росту у факультативних та облігатних психрофілів, при температурах які прямують до граничних для їх росту. Таким приладом є представлений графік Арреніуса граф. 1.

Графік 1. Крива Арреніуса для швидкості росту психрофілів
Графік дає уявлення, що швидкість росту для облігатного психрофілу залишається лінійною до -2 °С, а для факультативного відхилення починається за декілька градусів від нуля. Це вкрай важливе спостереження, оскільки воно може являтися фактором, що приводить до екологічного домінування в місце існуваннях з низькою температурою. Ці данні були підтвердженні в експериментах з морськими психрофілами в модельних системах, а також в умовах безперервного культивування. В цих дослідах облігатні психрофіли виявилися більш конкурентно спроможними, при температурах нижче 15 °С.
Іншим показником є температурний коефіцієнт росту (Q10 ) він визначається як відношення швидкостей при одній температурі та при іншій на 10 °С нижчій.
Q10 =Kt-/Kt
Формула 2. Температурний коефіцієнт росту
Для функції росту коефіцієнт може бути визначений в будь-якому інтервалі температур ΔТ з допомогою наступної формули:
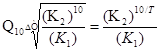
Формула 3. Температурний коефіцієнт Q10 для функції росту при температурі ΔТ
К1 та К2 –температурні коефіцієнту росту з різницею в 10 °С
Залежність швидкості біологічних реакцій від температури робить цей показник досить зручним чинником для порівняння мікроорганізмів по ступеню стійкості швидкості росту до зниження температури[12].
2.2 Фактори, що обумовлюють низьку максимальну температуру росту
Вважається, що верхня температурна межа росту мікроорганізмів залежить від термолабільних одного або кількох важливих у хімічному або структурному відношенні компонентів. Очевидно, якщо клітинний компонент (або будь-яка активність) відрізняється чутливістю до температури, близької до максимальної температури зростання, то між цією чутливістю і верхньою межею зростання повинен існувати причинний взаємозв'язок. Крім того, іноді вдається простежити позитивний взаємозв'язок між зменшенням термолабільності одного або декількох клітинних компонентів (або активності) і збільшенням максимальної температури зростання мікроорганізмів. Деякі дослідники культивували мікроорганізми при температурах як вище, так і нижче дійсної максимальної температури їх зростання. Дані, отримані в результаті таких експериментів, сприяють кращому розумінню механізмів, які визначають верхню межу зростання психрофільних і психротрофних мікроорганізмів. Синтез білків інтактними клітинами деяких психрофільних мікроорганізмів при відносно помірних температурах припиняється. Наприклад, синтез білків облігатним психрофілом Pseudomonas зупиняється при 22,5 ° С. Аналогічно в умовах безперервного культивування в хемостаті швидкість синтезу білків цим же організмом швидко падала при температурі близько 20 ° С. У цих умовах переставав синтезувати білки і одночасно втрачав життєздатність А4 cryophilus .
Слід зазначити, що температури, при яких відбувається денатурація рибосом психрофільних мікроорганізмів, набагато більша, ніж верхня межа зростання цих організмів. Разом з тим існує позитивна кореляція між температурою денатурації рибосом або рРНК і максимальною температурою зростання різних психрофілів. Таким чином, важко сказати чи має такий взаємозв'язок пряме відношення до верхньої температурної межі росту психрофільних мікроорганізмів. Характер фізичних змін рибосом під дією нагрівання не завжди знаходиться в прямій залежності з максимальною температурою зростання психрофільних мікроорганізмів. На відміну від дії підвищених температур на структуру рибосом на їх функціонування у психрофільних мікроорганізмів впливають температури набагато більш близькі до температур верхньої межі зростання. Рибосоми психрофілів можуть втратити активність при температурах, які мало відрізняються від максимальної температури росту мікроорганізму, хоча зростання при цьому ще можливе.
Вплив відносно невисоких температур може призводити до модифікації структурних компонентів мікроорганізмів, що ростуть при низьких температурах. Іноді відбуваються істотні зміни тонкої структури мікроорганізмів. В інших випадках спостерігаються фізіологічні порушення, викликані структурними змінами. Температура може також викликати ультраструктурні зміни у В.psychrophilus . Клітинні стінки цього психрофіла швидко руйнувалися при 40 ° С, що супроводжувалося злипання клітин та їх загибеллю. Модифікація структури клітин психрофілів під дією температури може позначатися і на їх зовнішньому вигляді. Крім безпосередньо спостережених або вимірюваних змін клітинної структури при дії відносно помірних температур з виділення внутрішньоклітинних речовин або справжньому повного лізису можна виявити модифікацію таких структур, як структури, відповідальні за проникність. Облігатний психрофіл Candida nivalis виділяє в навколишнє середовище різні внутрішньоклітинні розчинні сполуки: амінокислоти або короткі поліпептиди, неорганічний фосфат і нуклеотідмонофосфати. Виділення цих сполук не може бути викликано лізисом клітин, так як загальна кількість клітин, визначена прямим рахунком або за мутності, при цьому не зменшується. Далі, оскільки виділення з клітин білка і нуклеїнових кислот не виявлено, можна припускати, що клітинна мембрана піддається специфічній модифікації. Виділення названих вище сполук корелює із втратою клітинами життєздатності при 25 ° С; не виключено, що низька максимальна температура зростання цього мікроорганізму принаймні частково пояснюється термолабільністю клітинної мембрани.
Під дією відносно помірних температур може відбуватися лізис клітин психрофілів, а не тільки виділення з них з'єднань. Так, наприклад, цілі клітини В.psychrophilus зазнають лізис в калій-фосфатному буфері, рН 6,5 при температурах вище 28 °С. Чутливість ферментів деяких психрофільних мікроорганізмів до відносно помірних температур може бути причиною (чи однією з причин) того, що ці організми відрізняються низькою максимальною температурою росту. Зважаючи на це пояснення, слід, однак, враховувати ту обставину, що фермент може інактивувати під дією температури тільки у виділеному стані, але не в інтактних клітинах. Іноді цілісність клітини забезпечує повний або частковий захист активності термолабільних ферментів. Наприклад, малатдегідрогеназа в екстрактах клітин облігатного психрофіла V.marinus MP-1 легко інактивується при нагріванні. Активність цього ферменту знижувалася навіть при 0 ° С і майже повністю зникала в результаті інкубації протягом 10 хв при 30 ° С, тобто при температурі, що перевищує максимальну температуру росту. Аналогічну інактивацію зазнавав очищений в 20 разів препарат цього ферменту. Разом з тим, при температурах нижче оптимальної температури росту активність малатдегідрогенази в інтактних клітинах не змінювалася; отже, в цих температурних межах її захист забезпечується цілісністю клітин. Проте при температурах вищих оптимальної (15 °С) активність цієї дегідрогенази значно знижується навіть у інтактних клітинах даного психрофільного організму. Таким чином, чутливість цього ферменту до відносно низьких температур може служити причиною низького верхньої температурної межі зростання бактерій.