Реферат: Риниофиты. Бесполое размножение. Спорангии и спорогенез у древних высших растений
У Utota bruchii, например, величина спор, образующихся в одном и том же спорангии, сильно колеблется (от 21 до 32 (х). Однако связана ли эта разница в величине с половой дифференцировкой, остается невыясненным. Но для форм, близких к Ulota bruchii, из того же семейства Orthotriehaeeiae, именно для Macromitrium blumeivH Schlotheimia konigsbergii таковая дифференциревка была доказана М. Ф. Фдейшером. Споры у этик мхов также сильно разнятся в размера и, как показал М. Флейшер, яри ирорастании их из более крупных спор вырастают всегда карликовые мужские растения. Из мелких же спор развиваются нормальные женские особи.
М. Флейшер нашел также карликовые мужские и женские растения у Trismegistia brauniana (из Hypnobryales), хотя споры у этого мха внешне не отличаются друг от друга по размерам. Внутренняя половая дифференцировка, таким образом, у них имеется. Карликовые мужские растения Флейшер нашел и у многих других мхов.
Позднее гетероспория была обнаружена М. Эрнет-Шварценбах (Ernst-Schwarzenbach,1939) у ряда видов Macromitrium. Этим автором было исследовано 35 видов p. Macromitrium. Из них 16 видов оказались гомостюровыми, 14— явно гетероспоровыми, 5—неясно гетероспоровыми. Например у salakense микроспоры имеют в- среднем 19,6 ц в диаметре, мегаспоры — 36,7 ц. Микроспоры имеют желтую окраску и одеты гладкой оболочкой, мегаспоры — зеленые, оболочка их — бородавчатая. Отношение тех и других— 1 : 1. На выросшей из микроспор протонеме через 3—4 месяца после посева образовались карликовые растения с антеридиями. На протонеме, развившейся ив мегаспор, появились Стебля, но к моменту окончания опытов половых органов на них еще не было. На них, по-видимому, при дальнейшем развитии -должны были появиться архегонии.
Как видно из сказанного, между данными М. Флейшера и М. Эрнст-Шварценбах имеется известное противоречие. По М. Флейшеру, микроспоры являются женскими спорами, мегаспоры — мужскими, по М. Эрнст-Шварценбах, наоборот, то есть мхи ведут себя в этом отношении так же, как папоротникообразные. Вопрос о гегероспории у мхов требует, следовательно, дальнейшего исследования. Однако и в настоящий момент мы можем констатировать, что у лиственных мхов гетероспория в той или иной форме существует. Указаний на гетероепорию у печеночников нет.
Большинство гомоепоровых папоротникообразных обладает обоеполыми заростками. Это указывает на то, что в спорах их нет половой дифференцировки и все споры и внешне и внутренне вполне равноценны. Однако зачатки гегероспории мы можем уловить и у них. Так, по свидетельству К. Гебеля, споры типичного папоротника Dennstedtia pimctilobula при посеве дают однополые заростки, то есть часть их развивается в мужские, часть в женские заростки. То же самое наблюдается и у некоторых хвощей, как например, у Equisetum limosum, E. hiemale, E. silvestre. Споры стих. хвощей дают мужские и женские заростка, отличающиеся друг от друга размерами, строением и даже окраской. Мужские заростки — меньшей величины, более - простого строения по сравнению с женскими и более светлой, желтоватой окраски. И у Dennstedtia, и у названных выше хвощей пол споры, возможно, определяется генотипически, так что если все споры -по величине и форме одинаковы, то внутреннее их различие — весьма резкое. У других хвощей, как у Е. debile, E. laevigatum или Е. arvense, заростки — обоеполые, причем архегонии на них развиваются раньше антеридиев. Но наряду с обоеполыми у этих хвощей образуются и чисто мужские заростки; количество их весьма зависит от тех условий, при которых прорастают споры и развиваются заростки.
В этом отношении подробно -изучен Е. arvense E. Шратдем (Schratz, 1928). Названный автор исследовал, насколько внешние условия определяют у Е. arvense пол заростка. Результаты его исследования сводятся к следующему. При неблагоприятных условиях, например при очень густом посеве большинство заростков становится чисто мужскими. Отсюда можно сделать вывод, что все споры могут дать мужские заростки и что пол заростков определяется внешними условиями. При благоприятных условиях получается не менее 50%-мужских заростков. Остальные — сначала являются женскими, впоследствии же становятся обоеполыми: на них появляются антеридии.
Переход женских заростков в обоеполые особенно хорошо происходит тогда, когда не бывает оплодотворения, и жизнь заростка тем самым затягивается. Ухудшая условия существования, можно заставить женские заростки образовать антеридии. Заставить же женские заростки, улучшая внешние условия, перейти к образованию архегониев не удается. Отношение мужских заростков к женским (соответственно обоеполым), как 1:1, -наблюдается и среди заростков, находимых в природе.
Вое это позволяет думать, что однодомность является нормальным распределением полов у Е. arvense; если не удается все заростки довести до .обоеполого состояния, то это зависит от того, что пол заростка -определяется очень рано, при развитии спор, и раз определившись, -он сохраняется уже и в дальнейшей жизни заростка, не изменяясь. Интересно отметить, что в спорангиях некоторых видов ископаемого рода Calomosta-chys, наблюдались споры различной величины, что позволяет допускать у них фенотипическое определение пола.
Таким образом, у одних хвощей (Е. arvense, E. debile в др.) половая дифференцировка спор еще не установилась, и определение их пола находится в сильной зависимости от внешних условий, у других же видов (Е. limosum, E. hiemale и др.) половая дифференцировка уже совершилась и можно говорить о мужских и женских спорах. Гетероспорией эту дифференцировку назвать еще нельзя, так как внешнего различия между опорами здесь нет. Поэтому многие исследователи называют ее гетероталлизмом.
Ф. Жуйе-Лавернь показал, что споры хвощей различаются по своему химическому составу. Применяя различные прижизненные окраски: нейтральрот, крезильблау, митиленовую синьку, а также обрабатывая споры хвощей осмиевой кислотой и некоторыми другими реактивами, он обнаружил среди спор хвощей две группы, -отличающиеся друг от друга величиной окислительно-восстановительного потенциала — гН: одни из них — группы А, — имеют более низкий гН, чем другие (группы В). Исходя из того факта, что семяпочки покрытосемянных ведут себя так, как споры группы А, а пыльцевые зерна, как опоры группы В, Ф. Жуйе-Лавернь делает заключение, что споры группы А являются женскими, а группы В—мужскими опорами. Однако Е. Шратц не подтвердил результатов Ф. Жуйе-Лавернь.
Гетероталлизм в дальнейшей эволюции влечет за собой адтероспорию: споры начинают различаться между собой не только то внутреннему строению, но и внешнему — по величине, по строению оболочек и т. д. При этом мужские споры всегда гораздо мельче и проще устроены, чем женские. Вместе с тем гетероспория всегда, как это показывает изучение разноспоровых форм, связана с редукцией гаметофита. Редукции особенно сильно подвергается мужской гаметофит. Меньшая величина микроспор и большая редукция мужского гаметофита стоит, по-видимому, в связи с кратковременностью существования и функционирования его, с коротким сроком, в который совершается половой акт. Для развития немногих сперматозоидов, образующихся из микроспоры, не требуется .большого количества питательных веществ и отпадает вместе с тем необходимость в вегетативной ткани заростка; Немногочисленность Сперматозоидов, даваемых каждой микроопорой, восполняется большим числом микроспор, так что .общее количество сперматозоидов, образуемых особью, громадно и вполне обеспечивает половой процесс. Сильную редукцию мужского гаметофита мы видим у всех разноспоровых папоротникообразных. У оелагинелл, Isoetes, и водных папоротников вегетативная часть мужского заростка сводится, в сущности, к двум клеткам—ризоидальной и базальтом. Кроме них мужской заросток содержит один (Isoetes) или два антеридия (оелагинелли, водные папоротники), дающие немногие сперматозоиды. В еще большей степени редуцирован мужской гаметофит у голосемянных и покрытосемянных. У голосемянных при прорастании микроспор образуется одна (Cyoadales) или две Вегетативные клетки мужского заростка (Ginkgo, хвойные). Они очень скоро разрушаются и ко времени образования половых элементов от них не остается ни следа. Остальные клетки мужского заростка (клетка-ножка, клетка, дающая пыльцевую трубку) надо считать принадлежащими к антеридию.
У покрытосемянных же стерильных клеток мужского заростка совсем не образуется, имеется лишь клетка, вырастающая в пыльцевую трубку; ее следует считать принадлежащей антеридию. Но самый микроспорангий при этом не подвергся никакой особой эволюции, сохраняв свой характер или лептоспорангия (водные папоротники), или евспорангия (селагинелли, голо- и покрытосемянные).
Мегаспоры (женские споры) много крупнее микроспор и имеют более сложное строение, обладая толстыми, нередко многослойными оболочками со сложной структурой. У папоротникообразных, кроме того, в мегаспорах откладываются в большем или меньшем количестве запасные питательные вещества, за счет которых развивается женский заросток и зародыш на первых стадиях своей жизни. У голосеменные, у которых мегаспора не покидает мегаспорангия, питание развивающемуся заростку и зародышу доставляет спорофит.:
Женский заросток редуцирован значительно меньше, чем мужской. Он почти всегда представляет собой тканевое образование, и связан с оболочкой мегаспоры. Женский заросток или имеет вид небольшой зеленой пластинки (Salvinia, Azolla) или образует комплекс тонкостенных, плотно соединенных между собой клеток, заполняющих полость мегаспоры (селагинелла, Isoetes, голосемянные). Наибольшей редукции подвергся женский заросток у Marsiliaceae, где он сводится к двум-трем клеткам и одному архегонию.
Самый мегаспорагий у папоротникообразных не претерпел сложной эволюции и сохранил характер спорангия (ев спорангия или лептоспорэнгия), свойственный той или иной их группе. У семенных же растений (голо- или покрытосемянные) мегаспорангия превратился в семяпочку, пройдя сложную эволюцию в связи с тем, что мегаспора стала развиваться внутри мегаспорангия на Материнском растении и тем, что мегаспорангий был Образованием, из которого возникло ceмя. С семяпочками мы впервые встречаемся у семенных папоротников.
Во всех отделах голосемянных растений семяпочки построены в общем по одному плану. Семяпочка состоит из шуцеялуса, образующего центральную часть ее. Внутри нуцеллуса помещается эндосперм, построенный из тонкостенных, плотно соединенных между собой клеток. В эндосперме находятся архегогии. Снаружи нуцеллус одет покровом или интегументом, имеющим на вершине микропиле или семявход. -
Еще исследованиями В. Гофмейстера (1851) доказано, что нуцеллус морфологически представляет собой мегаспорам, а эндосперм — женский заросток, который развился внутри мегаспорангия из мегаспоры. Эта последняя здесь не высеивается из мегаспорангия, а остается внутри его, тут же про растает в заросток, который всю жизнь связан с спорофитом паразитируя, так сказать, на нем.:
Что же касается интегумента, то он является новым образованием, не имеющим гомолога у папоротникообразных. Но если ясна морфологическая природа нуцеллуса и эндосперма, то нельзя того же сказать про интегумент. Морфологическое значение и происхождение покрова остается неясным, и вполне возможно, что интегументы в разных порядках голосемянных растений не гомологичны.
В. Циммермавн приписывает интегументу листовое происхождение, думая, что он возник из сросшихся между собой в виде чашечки филлоидов. В пользу этою говорит, по его мнению, то, что у Lyginopteris, например интегумент не срастается с нуцеллусом, а свободно облегает его; далее у некоторых птеридоспермов, в особенности у Physostoma elegans, сосудистые пучки, пронизывающие интегумент, свободно оканчиваются в нем так же, как это наблюдается в чашечке. Однако отношение этих филлоидов к спорофиллам остается неясным.
Р. Пильгер (1927) .по поводу интегументов цикадовых говорит, что многое указывает на то, что толстый интегумент, свойственный этой группе, произошел из отдельных, сросшихся по длине частей, морфологическая природа которых неясна. Так, у Macrozamiia после удаления внешнего, сочного слоя интегумента можно видеть, что мииропиле окружено 8—И ребрами, разделенными короткими, глубокими ложбинками; у Geratoaamia и Encephalartos по каменистому слою семени проходят от основания его до микропиле хорошо выраженные ребра. В пользу того же предположения говорит и строение семян некоторых семенных папоротников, как например Physostoma elegans, упомянутой выше. Семена ее достигают 6 мм в длину и 2 мм в поперечнике. По оболочке вдоль их проходят 10 ребер, переходящие в свободные лопасти, окружающие венчиком верхушку нуцеллуса. Они играют роль микропилярной трубочки. Сосудистый пучок, войдя в халазу, дает десять ветвей, из которых каждая входит в ребро и продолжается в лопасть. Интегумент и нуцеллус срастаются почти до того места, где начинаются свободные лопасти покрова.
Оригинальный взгляд на происхождение интегумента голосемянных был высказан М. Бенсон (Benson, 1904). На основании изучения семян Telangium, она пришла к выводу, что семяпочка семенных папоротников возникла из еинангия. Произошло это таким образом, что расположенный в центре еинангия мегаспор авгий сохранил свою функцию и превратился в нуцеллус. Окружающие же его мегаспорангии стерилизовались и, срастаясь между собой, образовали интегумент. Этот последний является, таким образом, гомологом мегаопорангиев, а в конечном счете плодущих теломов В. Циммерманна.
К аналогичному выводу пришел С. Уольтон (Walton, 1953). Он на основании сравнительно морфологического анализа семян Pteridospermae считает, что интегумент и плюска семенных папоротников произошли из некоторого числа листовых образований путем их срастания между собой и обрастания ими нуцеллуса.
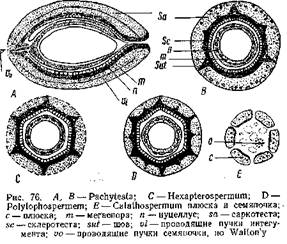
При этом интересно отметить, что число этик образований и в группе Lagenostomales и Trigonocarpales обычно кратно трем. У Calathospermqm плюсна состоит из 6 отдельных частей, окружающих мегаспорангии. У Hexapterospermum и Polylophospermum части эти срастаются и образуют общий покров, достигающий значительной толщины и дифференцирующийся натри Слоя — наружный мясистый (sarcotesta), второй — каменистый (solerotesta) и внутренний — тонкостенный. В местах сращения на склерютесте возникли ребра, вдающиеся в саркотесту (рис. 76). Этот покров семян Trigonacarpales соответствует плюске- Lagenostoma, а собственно интегумент, хорошо выраженный у Lagenostoma и свободно лежащий в плюске, в семенах Trigonocarpus срастается с сильно редуцированным нуцеллусом, образуя то, что называют у него нуцеллусом. И у Trigonocarpus и у Lagenostoma в семени находится только один мегаспорангии. Таким образом, Trigonocarpus по сравнению с Lagenostoma отличается более простым строением, и оба он более просты по сравнению с Caiiathospermum, у которого в купле находится несколько мегаслорангиев. Эволюция в семенах Pteridospermae шла, по мнению С. Уольтона, в направлении редукции числа мегаепорамгиев и упрощения строения их. Это упрощение выражается и в устройстве вершины нуцеллуса, довольно "сложного у Lagenostoma.
Выше изложенные соображения о возникновении интегумента у цикадовых могут быть так или иначе приложены и к хвойным. Однако в последнее время О. Гагеруп (Hagerup, 1933) выдвинул новое толкование интегумента у хвойных. На основании весьма подробного изучения истории развития женских шишек у ряда хвойных различных семейств он пришел к выводу, что интегумент представляет собой мегаепорофилл, несущий мегаопорангий, то есть пуцеллус с заключающейся в нем мегаспорой, а впоследствии — женским заростком. Это заключение подтверждают и явления позеленения семяпочек, наблюдаемые на женских шишках у многих хвойных. В таких случаях на месте семяпочек появляется вегетативный" лист (хвоя). Кроме того, можно встретить и переходы между хвоей в покровом семяпочки.
Напомним, что к такому же толкованию интегумента у кордаитов и хвойных пришел Р. Флорин (1951), на основания изучения ископаемых голосемянных. Речь об этом была выше, когда мы разбирали морфологию женских шишек у кордаитов и хвойных. У них, по толкованию Р. Флорина, интегумент является гомологом двух конечных лопастей мегаспорофишла, между которыми помещался мегагаюрангий. Но у тиссовых интегумент возник путем срастания двух или нескольких чешуи, соответствующих по своему положению мегаспорофиллам.
Неясным остается и Происхождение покровов у покрытосемянных растений. История развития не дает нам никаких указаний на филогению интегумента. По данным же сравнительной морфологии, согласно воззрениям одних авторов, они стеблевого (осевого) происхождения, по взглядам других — листового.
В пользу последнего взгляда, развиваемого, между прочим, А. Кернером (1900) говорят, прежде всего, явления озеленения семяпочек — превращения в листообразный орган внешнего интегумента. А. Кернер в позеленевших цветках Primula japonica нашел семяпочки, у которых внешний покров принял форму листа. Па нем помещался бугорок, зачаток нуцеллуса, а вокруг этого рудиментарного нуцеллуса находился кольцеобразный выроет, соответствующий внутреннему покрову. То же самое наблюдалось и у Trifolium repens, Rumex scutatus. Здесь также внешний покров превращается в лист, на котором сидит рудимент нуцеллуса с рудиментарным же внутренним покровом. И. Лотси у Hesperis находил семяпочки с превращенным в лист наружным покровом: на жилках его помещались зачаточные внутренние интегументы.
Эти факты подтверждают, листовое происхождение внешнего покрова. Но если это так, то внешний покров у покрытосемянных представляет собой мегасворофилл, несущий мегаопорангий (нуцеллус). Внутренний же интегумент —новообразование, вырост мегаспор ангия. Не будучи .генетически связан с листом, он листовидной формы при антолизе не принимает. В случае одною покрова семяпочки, можно принять, что он гомологичен мегаепорофиллу. Внутренний же покров или не развился или редуцировал. Но возможно, что произошло сращени