Реферат: Риниофиты. Бесполое размножение. Спорангии и спорогенез у древних высших растений
Риниофиты (Rhyniophyta), псилофиты (Psilophyta), самая древняя и примитивная вымершая группа (отдел) высших растений. Характеризовались верхушечным расположением спорангиев и равноспоровостью, отсутствием корней и листьев, дихотомическим или дихоподиальным (псевдомоноподиальным) ветвлением, примитивным анатомическим строением. Проводящая система — типичная протостела. Протоксилема располагалась в центре ксилемы; метаксилема состояла из трахеид с кольчатыми или (реже) лестничными утолщениями. Опорные ткани отсутствовали. Р. ещё не обладали способностью ко вторичному росту (меристемы у них были только верхушечные). Спорангии примитивные, от шаровидных (диаметром около 1 мм) до продолговато-цилиндрических (длиной до 12 мм), толстостенные. Гаметофиты. достоверно не известны (некоторые авторы считают гаметофитами горизонтальные корневищеподобные органы — так называемые ризомоиды). Риниофиты. произрастали на влажных и болотистых местах, а также в прибрежном мелководье. Отдел Риниофиты. включает один класс — риниопсиды (Rhyniopsida) с двумя порядками — Rhyniales (семества Cooksoniaceae, Rhyniaceae, Hedeiaceae) и Psilophytales (семейство Psilophytaceae). Для порядка Rhyniales характерны дихотомическое ветвление и тонкая, слабо развитая стела. Ксилема из трахеид с кольчатыми утолщениями. Древнейший представитель Риниофит. — род куксония, первоначально обнаруженный в Уэльсе в отложениях конца силурийского периода (около 400 млн. лет назад). Наиболее полно изучены нижнедевонские роды — риния и отчасти хорнеофит, у которого ризомоид (вверх от него отходили стебли, вниз — многочисленные ризоиды) был расчленён на чётковидно расположенные клубневидные сегменты, лишён проводящих тканей и целиком состоял из паренхимных клеток. Полагают, что в процессе эволюции ризомоиды. дали начало корням. У обоих родов стенка спорангия была многослойной, покрытой кутикулой. Хорнеофит характеризуется своеобразной спороносной полостью, которая образует купол, сводообразно покрывающий центральную колонку стерильной ткани, представляющей собой продолжение флоэмы стебля. Этим хорнеофит напоминает современный сфагнум. В семейства риниевых включают также род тениокрада, многие виды которого образовывали подводные заросли в среднем и верхнем девоне. В отдельное семейства хедеевых иногда выделяют нижнедевонские роды хедея и яравия. Нижнедевонский род сциадофит, обычно выделяемый в отдельное семейства сциадофитовых, — небольшое растение, состоявшее из розетки простых или слабо дихотомированных тонких стеблей со стелой. Для порядка Psilophytales характерны дихоподиальное ветвление и более сильно развитая стела. У наиболее известного рода — псилофит (из нижнедевонских отложений в Восточной Канаде) — неравно развитые ветви образовывали ложную главную ось дихоподия с более тонкими боковыми ветвями: стебель был окружен кутинизированной эпидермой с устьицами; поверхность стебля была голая или покрыта шипами длиной 2—2,5 мм, концы которых дисковидно расширялись, что, вероятно, указывало на их секреторную роль. Спорангии раскрывались продольной трещиной. К псилофиту близки нижнедевонские роды тримерофит и пертика.
Бесполое размножение. Спорангии
В предыдущей главе мы попытались проследить эволюцию спорангиеносных частей у архегоняальных растений. Теперь перейдем к рассмотрению самого процесса размножения. Попытаемся уловить основные моменты эволюции в развитии спорангиев и спор.
Как известно, у папоротникообразных существует два типа развития спорангия: евспорангиатный и лептоспорангиатный. При развитии спорангия по евепорангиатному типу в образовании его участвует целый участок ткани или во всяком случае группа клеток. Самый же спорангий в большинстве случаев отличается крупными размерами и толстой, многослойной стенкой.
Лептосиорангиатный тип характеризуется тем, что спорангий возникает из одной, эпидермической клетки, имеет небольшую величину и тонкую, однослойную стенку, снабженную более или менее ясно выраженным кольцом. По евепорангиатному типу развиваются спорангии у Psilotales, плаунов, хвощей, ужовников, маратциевых, у всех голосемянных. По этому же типу, вероятно, развивались спорангии у ископаемых представителей Lycopsida и Sphenopsida.
По лептоспорангиагному типу развиваются спорангии у настоящих папоротников. Примером евшорангиатного типа может служить развитие спорангия плауна — L. clavatura (рис. 69). По исследованиям П. А. Баранова (1924) спорангий L. clavatum закладывается на верхней стороне спорофилла, ближе к его основанию. Начало ему дает группа поверхностных клеток, которые уже на очень ранних стадиях несколько отличаются своим содержимым от соседних. Клетки эти делятся стенками, параллельными поверхности спорофилла, на наружные и внутренние. Из внутренних в дальнейшем образуется археопорий, из наружных — стенки спорангия. И те и другие клетки энергично делятся, в результате чего образуется бугорок. В дальнейшем в этом бугорке дифференцируется археспорий и двуслойная стенка спорангия. Клетки археспория отличаются более крупными размерами, крупными ядрами и густой протоплазмой. Между археспорием и стенкой спорангия образуется выстилающий слой — тапетум. Он отделяется от внутреннего слоя стенки спорангия, а в нижней части спорангия дифференцируется из прилежащей к археспорию ткани. Зачаток спорангия растет неравномерно: нижняя часть его отстает от верхней, в из нее образуется ножка спорангия. Клетки археспория обособляются и превращаются в материнские клетки спор, которые обычным путем дают тетрады и опоры.
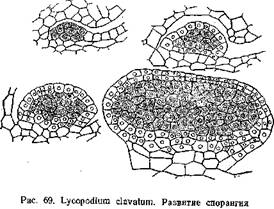
Также в общих чертах развиваются спорангии у селагинелл, хвощей и поилотов. Эволюция спорангиев выразилась в выработке приспособления для раскрывания спорангия и в возникновении особого Слоя, играющего роль в питании развивающихся спор и образовании у них сложных оболочек — тапетума или выстилающего слоя. Задачу раскрывания спорангия принимает на себя Наружный слой его стенки, превращаясь в так называемый экзотеций. Стенки его клеток утолщаются, причем утолщения эти у различных представителен папоротникообразных носят различный характер (балки, спирали и т. д.). Среди утолщенных клеток проходят ряды клеток снеутолщенными клетками (stomium). По ним проходит трещина, разрывающая стенку спорангия.
Выстилающий слой располагается между стенкой спорангия и спорогенным комплексом. Он отчленяется или от стенки спорангия (Lyoopodium) или от самого спорогенного комплекса, то есть наружные Слои этого последнего превращаются в тапетум (Ophioglossum, Equisetum). При этом различают два рода тапетума: секреторный и периплазмодиальный.
У секреторного тапетума стенки клеток сохраняются до конца его существования, и тапетум все время представляет собой слой клеток, облекающий внутреннюю поверхность спорангия. Но нередко клетки тапетума увеличиваются в размерах. При этом изменяется и характер их содержимого. Считают, что клетки секреторного тапетума выделяют теили иные вещества, которые участвуют в образовании спор и покрывающих их оболочек. Однако более подробно роль эта остается невыясненной.
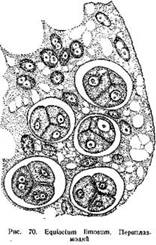
В периплаэмодиальном тапетуме стенки его клеток, иногда уже на ранних стадиях развития спорангия, расплываются к их протопласты сливаются вместе в одну сплошную массу, имеющую характер многоядерного плазмодия: его называют периплазмодием. После обособления материнских клеток спор, периплазмодий проникает в промежутки между ними, так что материнские клетки оказываются погруженными в плазматическую массу его (рис. 70). По мере развития спор и образования на них сложных оболочек периплазмодий становится все менее густым, в нем появляется много вакуолей. В конце концов, ко Времени вскрывания спорангия, периплазмодий исчезает.
Секреторный тапетум имеется у плаунов, селагинелл, у голосемянных; периплазмодиальный — у хвощей и ужовников. У псилотов также наблюдается периплазмодий, но у них он развивается своеобразно — не из тапетума, а из спорогенного комплекса. В этом последнем, на известной стадии развития спорангия наступает дифференцировка на плодущие и бесплодные клетки. Первые дают начало материнским клеткам спор и самим спорам, вторые же, после того, как разделяющие их стенки расплывутся, сливаются вместе и образуют периплазмодий.
Иной путь эволюции спорангиев выражается в слияния их между собой в синавгии. Синангии характерны для маратциевых папоротников (пор. Marattiales). При этом одни роды, как например, Angiopteris ли Archangiopteris, имеют еще отдельные спорангии, сидящие близко друг к другу. У других, как у Danaea, Christensenia или Marattia, спорангии настолько тесно срастаются, что границы отдельных, спорангиев в общем -синангии уловить нельзя. Открывается каждый спорангий отдельно продольной трещиной или круглым отверстием в общую борозду, проходящую между двумя рядами спорангиев (Marattia) или в углубление в центре синангия наблюдается и в других группах архегониат. Так оближенно сидят они, отчасти срастаясь, у Ophioglossales. У Cycas микроспоранши располагаются на нижней поверхности микро-спорофилла тесными группами. Но в наиболее, пожалуй, совершенной форме синангии представлены в ископаемой группе Whittleseyinae, относимой к Pteridospermae. У них (Christensenia) или, наконец, прямо на поверхность синангия (Danaea). Закладываются спорангии у Angiopteris отдельно, развиваясь из группы клеток У Marattia спорангии закладываются в общей плаценте, появляющейся в виде двух параллельных валиков, располагающихся вдоль жилки на нижней поверхности листа. Начало им дает целая группа клеток, которую, однако, по исследованиям Ф. Боуэра, можно свести к одной клетке.
Тенденция сливаться вместе и образовывать синангии срастаются микроопорангии, причем срастаются они настолько полно, что получается цельное тело, внешне напоминающее семя ли шлод.
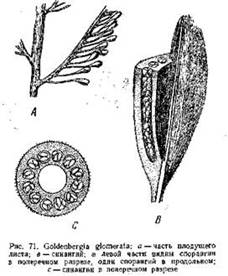
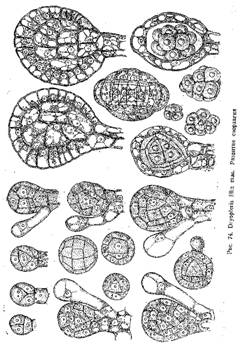
У Goldenbergia glomerata, например, (рис. 71) такой плод представлял собой овальное тело около 6—8 -мм длиной, сидящее на длинной тонкой ножке (10—15 мм). Внутри •его находилась обширная полость, а стенка была построена из вегетативной ткани, в толще которой заключалось 12—18 трубчатых спорангиев. Они располагались в один ряд вокруг центральной полости и содержали в себе довольно крупные микроспоры (0,3—0,4 мм). Внешняя поверхность плода была покрыта короткими шипиками.
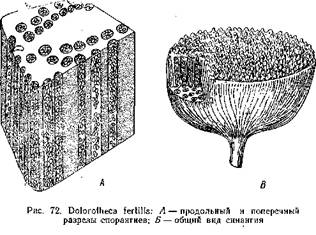
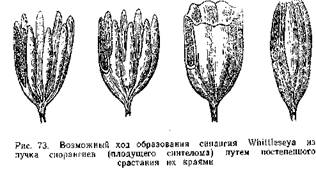
Подобное же строение имели и синангии Aulotheca. У Whittleseya elegans синангии имел форму бокала на тонкой ножке. Стенку бокала образуют сросшиеся между собой трубчатые спорангии с очень толстыми стенками. Dolorotheca мела чашевидные синангии, причем полость чаши была заполнена вегетативной тканью, в которую были вкраплены трубчатые спорангии (рис. 72). Т. Галле, подробно изучивший плодоношение Whittleseyinae, считает, что синангии их могли возникнуть путем тангентальвого сращения трубчатых спорангиев, подобно тому, как листья могли возникнуть ив дихотомически разветвленных теломов благодаря срастанию и с краями в одной плоскости.
Ход эволюции синангиев Whittleseyinae Т. Галле иллюстрирует следующей схемой (рис. 73). На ней первая фигура изображает гипотетическую исходную форму — плодущий синтелом. Наличие групп спорангиев Psilophyton, сидящих на концах теломов, подтверждает построение такой гипотетической формы. На следующей стадии спорангии срослись в нижней своей половине. Вершины их остаются свободными.
Это плодоношение типа Codonotheca. Далее срастание спорангиев идет до их вершин, получается бокаловидный синангии, как у Whittleseya elegans. Наконец, спорангии смыкаются и срастаются своими вершинами —образуется плод, полый внутри со спорангиями, находящимися в его стенке—Aulotheca, Goldenbergia.
Лептоспорангиатный тип развития спорангия характеризует типичные папоротники (пор. Filioales). Он отличается тем, что спорангий здесь возникает из одной эпидермической клетки, и сам спорангий имеет однослойную стенку и относительно малые размеры. Развитие его происходит по следующей схеме (рис. 74).
Клетка эпидермиса плаценты разрастается, выпячивается над общим уровнем эпидермиса и отделяется поперечной перегородкой от остальной ткани плаценты. Эта отделившаяся клетка и дает начало всему спорангию. В ней появляется горизонтальная перегородка, разделяющая ее на 2 клетки: нижнюю — меньшую, плоскую и верхнюю — большую, выпуклую. Нижняя в дальнейшем образует ножку спорангия, из верхней же развивается самый спорангий. В верхней клетке появляются последовательно три косые перегородки, пересекающие друг друга под некоторым углом. Первая перегородка проходит почти вертикально под небольшим углом к горизонтальной стенке, вторая пересекает под острым углом первую, а третья (на рисунке невидимая) пересекает тоже под острым углом две первые. Эти перегородки разделяют одноклетный зачаток спорангия на четыре клетки: одна из них занимает верхушку его и имеет форму трехгранной пирамиды, обращенной вершиной вниз, а выпуклым основанием наружу; три остальные лежат ниже ее, примыкая к ее граням. Далее, в пирамидальной клетке образуется горизонтальная перегородка.
Теперь весь зачаток состоит из пяти клеток— одной пирамидальной, внутренней, и четырех наружных, окружающих ее. Внутренняя клетка является археспориальной клеткой, и в нее развивается содержимое спорангия — опоры и выстилающий слой. Наружные же дают начало стенке спорангия. Они делятся только радиальными перегородками, так что стенка спорангия все время остается однослойной. В археспориальной же клетке появляются перегородки, тангенгальные, параллельные ее граням. Отделившиеся таким образом четыре таблитчатые клетки дают начало тапетуму, внутренняя же является опорообразующей, из нее развиваются споры. Клетки тапетума снова делятся тангентальными перегородками, вследствие чего он становится двуслойным. В то же время делится и опорообразующая клетка, постепенно распадаясь на 16 клеток. Вскоре стенки клеток тапетума расплываются и протопласты их сливаются, образуя периплазмодий. Клетки же археспория обособляются друг от друга и округляются, превращаясь в материнские клетки опор, которые обычным путем дают начало спорам.
Евопорангиатный тип спорангиев встречается у наиболее древних и примитивных групп среди ныне живущих папоротникообразных. Он же характерен) и для ископаемых папоротникообразных. Это заставляет нас смотреть на евшорангий как на древний, примитивный тип спорангия, от которого уже произошел лептосггорангий, свойственный к тому же более молодым группам папоротников (Filices). Об этом говорит и то, что, как показывают сравнительные исследования, между евопорангиатный я лептоспорангиатным типами развития существует ряд постепенных переходов, связывающих их между собой. Вместе с тем эти переходы указывают на ход эволюции лептоспорангия.
На рис. 75 дана схема заложения опорангия у различных папоротников: рис. а относится к типичному лаптоспорангию (сем. Polypodiaeeae), а рис. g— к типичному евспорангию (Angiopteris). Между ними располагаются промежуточные типы, принадлежащие к различным родам. Понятно, эволюция лептоспорангия совершалась в направлении, обратном расположению рисунков. У Angiopteris археспорий закладывается глубоко под поверхностью и самая археспориальная клетка имеет форму куба: ограничивающие ее стенки пересекаются под прямым углом. У Todea (сем. Osmundaceae) наблюдаются два варианта развития опорангия. В одном случае ([) археспорий имеет в разрезе трапециевидную форму, благодаря тому, что антиклины, ограничивающие археспорий, наклонены под некоторым углом друг к другу. В другом случае (более мелкие спорангии) эти антиклины пересекаются, и археспорий принимает форму четырехгранной пирамиды, обращенной вершиной вниз, то есть форму, характерную для археспория лептоепорангия. И у Angiopteris и у Todea в образовании спорангия принимают участие и прилежащие к археспорию клетки. Развитие спорангия у Schiziaea (d), Alsophila(с) и Ceratopteris (Ь) представляет переход к типичному лептоспорангиатному типу (а). Археспорий окончательно принимает форму четырехгранной пирамиды и все меньшее число эпидермических клеток идет на развитие спорангиев. У Polypo-diaceae (а) весь спорангий развивается из одной эпидермической клетки.
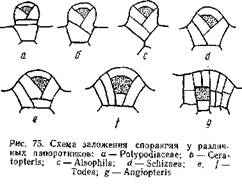
Таким образом, эволюция спорангия у папоротников шла в направлении редукции, которая и приведет к образованию лептоспорангия. Эволюция последнего, кроме того, выразилась в создании длинной и тонкой ножки, выработке кольца и в общем уменьшении количества производимых спорангием спор. В результате получился типичный лептоспорангий малой величины с длинной ножкой и однорядным кольцом. Число спор, образующихся в спорангии, невелико, но число спорангиев сильно возрастает, благодаря чему общее количество спор, производимых растением, не только не уменьшается, но и весьма увеличивается.
--> ЧИТАТЬ ПОЛНОСТЬЮ <--